Nessuno può dubitare dell’impegno di Dawkins nel difendere la teoria evoluzionistica dai Negazionisti (Creazionisti e sostenitori dell’Intelligent Design). Benché sottesa da una grande passione per la conoscenza scientifica e da una vasta cultura in ambito naturalistico, la sua opera riconosce come limite la tendenza ad assumere L’origine delle specie come una sorta di Bibbia laica, che va rispettata alla lettera.
Ciò significa non solo insistere sul fatto che la teoria evoluzionistica non è, come sostengono i Negazionisti, una congettura ancora lungi dall’essere dimostrata:
“L'evoluzione è un fatto. Oltre ogni ragionevole dubbio, oltre ogni serio, sano, informato, intelligente dubbio l'evoluzione è un fatto. Le sue prove sono consistenti almeno quanto quelle dell'Olocausto, che pure dispone di testimoni oculari. E’ la pura verità che siamo cugini degli scimpanzé, cugini più lontani delle scimmie non antropomorfe, cugini ancora più lontani degli orittèropi e dei lamantini, cugini ancora più lontani delle banane e delle rape, e via elencando. Non è una verità che salta agli occhi. Non è una tautologia, una verità evidente e lapalissiana, e in passato la maggior parte delle persone, anche di quelle colte, non la riteneva tale. Non salta agli occhi, ma è la verità. Lo sappiamo perché è confermata da una valanga di prove. L'evoluzione è un fatto, e questo libro lo dimostrerà. Nessuno scienziato rispettabile la confuta e nessun lettore obiettivo chiuderà il libro dubitandone.” (pp. 11-12)
La mole del libro, di ben 370 pagine, di fatto è dovuta alla “valanga” di prove che Dawkins adduce ed analizza fin nei particolari.
Il problema è che egli intende dimostrare anche la validità dei presupposti “ideologici” - il gradualismo e l’adattamentismo - che Darwin ha ritenuto necessari al fine di supportare l’ipotesi della selezione naturale. Da tempo ormai un numero crescente di biologi evoluzionisti ha messo in discussione quei presupposti ritenendoli il punto debole del darwinismo e del neodarwinismo, in quanto essi non si accordano con il “silenzio fossile” che riguarda lunghi periodi dell’evoluzione, con le prove di periodiche fasi caratterizzate viceversa dalla comparsa, in tempi geologici relativamente brevi, di un numero rilevante di nuove specie, e con la presenza, lungo la scala dei viventi, di strutture e funzioni che non sembrano avere un significato originariamente adattivo, quindi non possono essere state selezionate.
La teoria degli equilibri punteggiati, messa a fuoco da S. Y. Gould e N. Edredge, si è fatta carico di questi problemi dimostrando che è possibile integrarli con il darwinismo a patto di sormontare il principio gradualista e l’ideologia iperadattamentista. Né Gould né Edredge hanno mai negato il valore della selezione naturale. Essi, però, hanno tenuto conto del pensiero di Darwin, che riteneva essere questa il principale ma non l’unico meccanismo dell’evoluzione naturale.
Dawkins, viceversa, insiste nel ritenere la selezione naturale come unico meccanismo, ed è costretto pertanto a convalidare anche il gradualismo e l’adattamentismo, vale a dire a pensare che l'evoluzione delle forme viventi sia avvenuta lentissimamente nel corso del tempo sulla base di impercettibili variazioni strutturali, dovute a mutazioni genetiche, esitate infine nella nascita di nuove specie. L'unica novità teorica che egli accetta è la deriva genetica, cioè la possibilità che la diffusione di geni mutanti in un a popolazione possa avvenire più facilmente laddove essa giunge a trovarsi in una situazione di segregazione ambientale.
Egli, insomma, si può considerare un ultradarwinista o - ed è la stessa cosa - il più accanito sostenitore del neo-darwinismo, il cui paradigma ha addirittura radicalizzato, assumendo il gene (e non l’individuo) come unità dell’evoluzione.
Il saggio realizza un intento e ne fallisce un altro: dimostra in maniera inconfutabile che la teoria darwiniana dispone di prove molteplici e schiaccianti a suo favore, sicché metterla in discussione o ritenerla una congettura appare francamente ridicolo. Non dimostra, invece, la fondatezza né del gradualismo né dell’iperadattamentismo.
Per rendersi conto di quest’ultimo aspetto occorre leggere il capitolo VI e il VII, dedicati rispettivamente agli anelli mancanti nella scala evolutiva e nell’antropogenesi. Li riporto integralmente per poi commentarli.
I creazionisti sono profondamente innamorati della documentazione fossile, perché hanno imparato (gli uni dagli altri) a ripetere continuamente, come un mantra, che essa è piena di «lacune» lanciando la sfida: «Mostrateci gli stadi "intermedi"!». Con gioia (con grande gioia) immaginano che queste «lacune» mettano in imbarazzo gli evoluzionisti. In realtà, se è già una fortuna avere dei fossili, è una fortuna sfacciata avere a disposizione la folta massa di reperti che documentano la storia evolutiva e che in molti casi costituiscono, a tutti gli effetti, ottimi «intermedi». Nei capitoli IX e X dimostrerò che non abbiamo bisogno di fossili per dimostrare che l'evoluzione è un fatto. L'evoluzione sarebbe assolutamente certa anche se non si fosse mai fossilizzato neanche un cadavere. E’ una fortuna in più che abbiamo davvero ricche riserve di fossili e che ogni giorno ne scopriamo di nuovi. Per molti dei principali gruppi animali, le prove fossili dell'evoluzione sono straordinariamente esaurienti. Tuttavia vi sono, com'è naturale, delle lacune e i creazionisti le amano alla follia.
Torniamo all'analogia del detective che arriva sulla scena di un delitto del quale non ci sono stati testimoni oculari. Hanno sparato al baronetto. Le impronte digitali, le orme, il DNA a cui si è risaliti da una macchia di sudore sulla pistola e un forte movente fanno tutti pensare al maggiordomo. E' un caso semplicissimo: la giuria e tutti i presenti in aula sono convinti che sia stato lui. Ma all'ultimo momento, proprio un attimo prima che la giuria si ritiri per discutere di quello che appare l'inevitabile verdetto di colpevolezza, viene scoperta una prova: qualcuno ricorda che il baronetto aveva installato telecamere spia contro i rapinatori. Col fiato sospeso, la corte guarda i filmati, uno dei quali mostra il maggiordomo nell'atto di aprire un cassetto nella sua stanza, estrarre una pistola, caricarla e uscire con aria furtiva e una luce malvagia negli occhi. Parrebbe un'ulteriore conferma dei sospetti nei suoi confronti, ma bisogna guardare anche il seguito. L'avvocato del maggiordomo osserva astutamente che non c'erano telecamere nascoste nella biblioteca dove è avvenuto il delitto, né ce n'erano nel corridoio che porta dalla stanza del maggiordomo alla biblioteca. Agitando l'indice nel tipico, suggestivo gesto da avvocato, egli proclama: «C'è una lacuna nel filmato. Non sappiamo che cosa è successo dopo che il maggiordomo ha lasciato la sua camera. E’ evidente che le prove non sono sufficienti a condannare il mio cliente».
Invano il pubblico ministero osserva che c'era una seconda telecamera nella sala da biliardo e che il relativo filmato mostra, attraverso la porta aperta, il maggiordomo avanzare in punta di piedi, con la pistola in pugno, lungo il corridoio che porta alla biblioteca. Il nuovo tassello non dovrebbe colmare la lacuna, non dovrebbe dimostrare che prove schiaccianti ormai inchiodano il maggiordomo? No. Trionfalmente, l'avvocato della difesa gioca il suo asso. «Non sappiamo che cosa è successo prima e dopo che il maggiordomo è passato davanti alla porta aperta della sala da biliardo. Adesso vi sono due lacune nel filmato. Signore e signori della giuria, ho concluso: in questo momento vi sono ancora meno prove contro il mio cliente di quante ve ne fossero prima.»
Come le telecamere del racconto giallo, la documentazione fossile è un bonus, una circostanza fortuita in cui non pensavamo di avere la fortuna di incappare. Le prove per incriminare il maggiordomo erano più che sufficienti anche senza telecamere nascoste e la giuria stava per rilasciare un verdetto di colpevolezza già prima che si scoprisse il filmato. Analogamente, lo studio comparato delle specie moderne (capitolo X) e della loro distribuzione geografica (capitolo IX) ha prodotto prove più che sufficienti a dimostrare la realtà dell'evoluzione. Non abbiamo bisogno di fossili: l'evoluzione è inconfutabile anche senza di essi, sicché è paradossale usare le lacune nella documentazione fossile come prove contrarie all'evoluzione. Avere fossili è semplicemente una fortuna in più.
Una vera smentita dell'evoluzione, e molto convincente, sarebbe invece scoprire anche un solo fossile nello strato geologico sbagliato. Com'è noto, a chi gli chiedeva di nominare una prova che avrebbe dimostrato la falsità della teoria evoluzionistica, J.B.S. Haldane rispose: «Conigli fossili nel Precambriano!». Non sono mai stati rinvenuti né conigli né altri fossili anacronistici. Tutti i reperti che abbiamo trovato, e sono numerosissimi, si sono presentati senza una sola eccezione nella giusta sequenza temporale. Certo, esistono lacune là dove non si sono scoperti fossili di sorta, ed è perfettamente comprensibile che non se ne siano scoperti; ma non si è mai rinvenuto un solo reperto che risalisse a un periodo precedente quello in cui si sarebbe dovuto evolvere. E’ un fatto assai significativo (e che in base al creazionismo non avrebbe ragion d'essere). Come ho spiegato concisamente nel IV capitolo, una teoria valida, capace di rispettare tutti i crismi scientifici, è una teoria che si può confutare e che tuttavia non viene confutata. L'evoluzione potrebbe essere facilmente smentita se anche un solo fossile comparisse nell'ordine cronologico sbagliato. L'evoluzione ha superato questo test a pieni voti. Chiunque voglia confutarla vaghi tra le rocce alla scrupolosa quanto disperata ricerca di fossili anacronistici. Forse ne troverà uno. Scommettiamo di no?
La lacuna più grande, e anche la più amata dai creazionisti, è quella precedente la cosiddetta esplosione del Cambriano. Oltre mezzo miliardo di anni fa, nel periodo Cambriano, apparve «all'improvviso» la maggior parte dei grandi phyla, i principali tipi del mondo animale. «All'improvviso» non significa che il fenomeno sia stato subitaneo, ma che nessun fossile di quei gruppi è mai stato rinvenuto in rocce più antiche di quelle del Cambriano: il periodo di cui stiamo parlando dura infatti circa venti milioni di anni. Venti milioni di anni sembrano pochi quando si parla di un'epoca lontana cinquecento milioni di anni, ma naturalmente rappresentano lo stesso esatto arco di tempo evolutivo di venti milioni di anni odierni. In ogni caso, resta un fenomeno abbastanza «improvviso». Come ho scritto nell'Orologiaio cieco, nel Cambriano un notevole numero di grandi phyla animali sono
già in uno stato di evoluzione avanzato la primissima volta che compaiono tra i fossili. E un po' come se fossero apparsi all'improvviso, senza alcuna storia evoluzionistica. Non occorre dire che questo fatto ha deliziato i creazionisti.
L'ultima frase dimostra che già all'epoca ero abbastanza sveglio da capire quanto i creazionisti amassero l'esplosione del Cambriano. Allora però (era il 1986) non fui abbastanza sveglio da intuire che con gran disinvoltura avrebbero usato il mio brano come cavallo di battaglia per portare acqua al loro mulino, omettendo accuratamente le mie argomentazioni. Così, d'impulso, ho cercato nel Web la frase «è un po' come se fossero apparsi all'improvviso, senza alcuna storia evoluzionistica» e ho trovato non meno di 1250 occorrenze. Per controllare in maniera approssimativa se, come pensavo, la maggioranza delle voci fosse costituita da citazioni creazionistiche tendenziose, ho provato a cercare la frase immediatamente successiva del mio libro: «Gli evoluzionisti di ogni specie credono però che questa sia davvero una grande lacuna nella documentazione fossile», e ho ottenuto un totale di 63 occorrenze. Il rapporto tra 1250 e 63 è 19,8: potremmo chiamarlo l'«indice delle citazioni tendenziose».
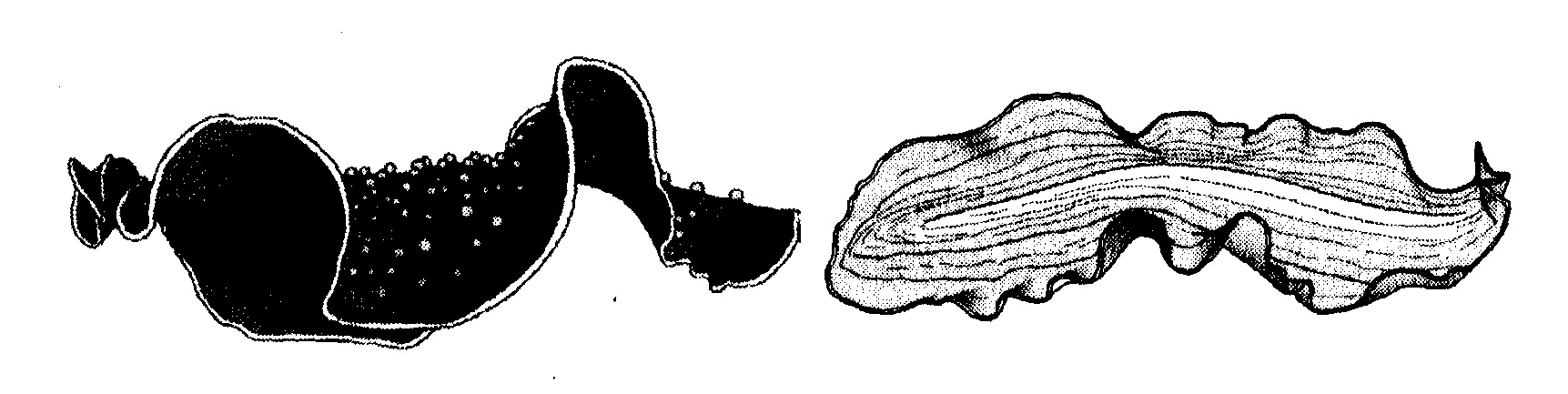
Ho analizzato estesamente l'esplosione del Cambriano, soprattutto nell'Arcobaleno della vita. Qui aggiungerò solo un concetto, illustrato dai platelminti. Questo grande phylum di vermi comprende parassiti come i trematodi e la tenia, che sono di grande importanza medica. I miei preferiti, però, non sono parassiti: si tratta dei turbellari, i quali, con oltre quattromila specie, sono numerosi quanto tutti i mammiferi messi assieme. Alcuni, come dimostrano i due della figura 6.1, sono creature meravigliose.
Sono diffusi sia in acqua sia sulla terraferma ed è probabile che lo siano stati per moltissimo tempo. Parrebbe quindi logico aspettarsi una loro ricca storia fossile; purtroppo, invece, è praticamente inesistente. A parte un pugno di ambigui icnofossili, non è stato rinvenuto un solo platelminta fossile. I platelminti, per essere vermi, sono «già in uno stato di evoluzione avanzato la primissima volta che compaiono tra i fossili. E un po' come se fossero apparsi all'improvviso, senza alcuna storia evoluzionistica». Ma in questo caso «la primissima volta che compaiono» non è il Cambriano, bensì oggi. Capite che significa o almeno dovrebbe significare per i creazionisti? I creazionisti credono che i platelminti siano stati creati la stessa settimana in cui furono creati tutti gli altri esseri viventi e che quindi abbiano avuto esattamente lo stesso tempo di tutti gli altri animali per fossilizzarsi. In tutti i secoli in cui tanti animali dotati di ossa o carapaci si fossilizzavano a migliaia, i platelminti vissero felicemente accanto a loro, senza però lasciare nelle rocce alcuna traccia rilevante della loro presenza. Allora perché scandalizzarsi tanto delle lacune nella documentazione degli animali che si fossilizzano, se la storia passata dei platelminti consiste in un'unica grande lacuna, benché, dal punto di vista creazionistico, essi siano vissuti quanto tutti gli altri animali? Se la lacuna precambriana viene addotta come prova del fatto che la maggior parte degli animali sia comparsa all'improvviso nel Cambriano, alla stessa identica «logica» si dovrebbe ricorrere per dimostrare che i platelminti hanno cominciato a esistere ieri. Invece i creazionisti affermano che i vermi piatti non sono nati ieri, bensì la stessa settimana in cui fu creato tutto il resto. O è vera una cosa o è vera l'altra: si escludono a vicenda. Questo ragionamento logico annulla completamente, di colpo, la pretesa dei creazionisti di usare la lacuna precambriana come prova contro l'evoluzione.
E gli evoluzionisti, come spiegano che vi siano così pochi fossili prima del Cambriano? E’ probabile che, quali che siano i fattori responsabili della mancanza di platelminti fossili in tutte le ere geologiche fino a oggi, essi abbiano influito anche sul resto del regno animale prima del Cambriano. Probabilmente la maggior parte degli animali precambriani era costituita da invertebrati come i moderni platelminti, magari piuttosto piccoli come i moderni turbellari e composti da un materiale poco adatto alla fossilizzazione. Poi, mezzo miliardo di anni fa, accadde qualcosa che permise agli animali di fossilizzarsi liberamente: per esempio la comparsa di scheletri duri e mineralizzati.
Un altro modo di indicare la «lacuna nella documentazione fossile» era l'espressione «anello mancante», che fu molto in voga nell'Inghilterra tardo‑vittoriana e persistette fino alla prima metà del XX secolo. Frutto di un fraintendimento della teoria darwiniana, era usata come insulto un po' come è usato oggi, colloquialmente (e ingiustamente), il termine «neandertal». Nell'elenco di citazioni che l'Oxford English Dictionary riporta per illustrare il significato dell'espressione, ce n'è una del 1930 che riguarda D.H. Lawrence. Una donna, racconta il romanziere, gli scrisse per dirgli che il suo nome «puzzava» e concludeva: «Tu, che sei un incrocio tra un anello mancante e uno scimpanzé».
Il significato originario dell'espressione, molto confuso come dimostrerò, era che alla teoria darwiniana mancasse il nesso fondamentale tra l'uomo e gli altri primati. L'oxford riporta un'altra citazione, risalente all'Ottocento: «Ho sentito parlare di un anello mancante tra uomini e puggies» (puggie era un termine dialettale scozzese per «scimmia»). Ancora oggi i negazionisti ripetono, con compiaciuto tono di dileggio: «Ma non avete ancora trovato l'anello mancante», e spesso aggiungono una frecciatina sull'Uomo di Piltdown. Non si sa chi abbia architettato la burla del Putdown, ma chiunque sia stato, deve rispondere di molti danni. (* In genere si sospetta sia stato il paleontologo Charles Dawson, ma Stephen Jay Gould ha avanzato la suggestiva ipotesi che fosse Pierre Teilhard de Chardin. Il lettore forse ricorderà che Teilhard era il teologo gesuita il cui ultimo libro, Il fenomeno umano, ricevette la stroncatura più tremenda di tutti i tempi dall'impareggiabile Peter Medawar (il quale la incluse poi in The Art of the Soluble e Pluto's Republic).
Il fatto che uno dei primi candidati alla qualifica di uomo‑scimmia fossile sia risultato essere una bufala ha fornito ai negazionisti il pretesto per ignorare i numerosissimi fossili autentici; e i creazionistì non smettono di andarci a nozze. Se solo guardassero i fatti, presto scoprirebbero che esiste ormai una ricca riserva di fossili intermedi i quali collegano l'Homo sapiens moderno con l'antenato comune che esso condivide con gli scimpanzé. Sul versante umano dello spartiacque, intendo. Perché, particolare interessante, non vi sono ancora fossili che collegano quell'antenato (né scimpanzé né uomo) agli scimpanzé moderni. Forse non ce ne sono perché gli scimpanzé vivono in foreste che non offrono condizioni adatte alla fossilizzazione. Semmai sarebbero gli scimpanzé, e non gli uomini, ad avere oggi il diritto di lamentarsi dell'anello mancante!
Uno dei significati dell'espressione «anello mancante» è dunque quello del presunto divario tra l'uomo e il resto del regno animale. Inteso in questo senso, l'anello mancante non manca più e dire che non manca è dire poco. Tornerò sull'argomento nel prossimo capitolo dedicato ai fossili umani.
Un altro significato è quello della presunta scarsità delle cosiddette «forme transizionali» tra i maggiori gruppi, per esempio tra rettili e uccelli o tra pesci e anfibi. «Mostrateci gli stadi intermedi!» Gli evoluzionisti spesso rispondono al grido disfida dei negazionisti lanciando loro le ossa dell'Archaeopteryx, il famoso «stadio intermedio» tra «rettili» e uccelli. Come dimostrerò, è un errore. Archaeopteryx non è la risposta alla sfida, perché non c'è sfida degna di risposta. Proporre come argomento un unico fossile famoso come Archaeopteryx asseconda la credenza erronea. In realtà, di un gran numero di fossili si può dire che siano stadi intermedi tra qualcosa e qualcos'altro. Il presunto problema cui Archaeopteryx parrebbe rispondere si basa su una concezione superata, quella della cosiddetta «Grande catena dell'essere". Affronterò l'argomento più avanti in questo stesso capitolo.
Di tutte le stupide critiche incentrate sull'«anello mancante», le più stupide sono due (o le loro numerose varianti). La prima è: «Se l'uomo è disceso dalla scimmia attraverso le rane e i pesci, perché la documentazione fossile non comprende le "ranimmie"?». Ho visto un creazionista islamico chiedere con aria truce come mai non ci fossero fossili di coccodranatre (ill. 8b). La seconda è: «Crederò all'evoluzione quando vedrò una scimmia partorire un bambino». Con quest'ultima dichiarazione si commette lo stesso errore che con quella precedente, e in più quello di pensare che i grossi cambiamenti evolutivi avvengano da un giorno all'altro.
Per caso, due di queste credenze erronee compaiono l'una accanto all'altra nel lungo elenco di commenti che sono seguiti a un articolo del «Sunday Times» di Londra su un documentario televisivo di argomento darwiniano che avevo presentato:
L'opinione di Dawkins sulla religione è assurda, perché la stessa evoluzione non è altro che una religione. Bisogna credere che proveniamo tutti da una singola cellula.., e che una lumaca possa diventare una scimmia ecc. Ah, ah, è la religione più ridicola che sia mai stata concepita! Joyce, Warwickshire, GB
Dawkins dovrebbe spiegare perché la scienza non è riuscita a trovare gli anelli mancanti. La fede in una scienza priva di fondamento è una favola ancora più incredibile della fede in Dio. Bob, Las Vegas, USA
In questo capitolo smonterò tutte queste credenze erronee, a cominciare dalla più stupida di tutte, dato che la risposta a essa servirà da introduzione alle altre.
«Perché la documentazione fossile non comprende le "ranimmie"?» Naturalmente le scimmie non discendono dalle rane: nessun evoluzionista sano di mente lo ha mai sostenuto, né ha mai sostenuto che le anatre discendessero dai coccodrilli o viceversa. Le scimmie e le rane condividono un antenato, che non somigliava affatto né alle prime né alle seconde. Forse somigliava un poco a una salamandra, e in effetti abbiamo dei fossili simili a salamandre che risalgono all'epoca giusta. Ma non è questo il punto.
Ognuna delle innumerevoli specie animali condivide un antenato con tutte le altre. Se si capisce così poco l'evoluzione da credere che sia logico trovare fossili di ranimmie o coccodranatre, si dovrebbe ironizzare anche sulla mancanza di un canepotamo e un elefanzé. Anzi, perché limitarsi ai mammiferi? Perché non pretendere che siano esistiti un cangurfaggio (intermedio tra canguro e scarafaggio) o un polpardo (intermedio tra un polpo e un leopardo)? Si possono coniare infiniti termini mettendo insieme un animale con l'altro. Naturalmente gli ippopotami non sono discesi dai cani, né i cani dagli ippopotami. Gli scimpanzé non sono discesi dagli elefanti (o viceversa), come le scimmie non sono discese dalle rane. Nessuna specie moderna è discesa da un'altra specie moderna (se tralasciamo le divisioni molto recenti). Come si trovano fossili che si avvicinano all'antenato comune di una rana o una scimmia, così si trovano fossili che si avvicinano all'antenato comune di elefanti e scimpanzé. Eccone per esempio uno che si chiamava Eomaia (fig. 6.2) e che visse all'inizio del Cretacico, poco più di 100 milioni di anni fa.

Come si può constatare, Eomaia non somigliava né a uno scimpanzé né a un elefante. Vagamente simile a un toporagno, somigliava forse abbastanza al loro antenato comune, del quale era all'incirca contemporaneo; come si può vedere, si sono verificati molti cambiamenti evolutivi sia nel percorso che va dall'antenato affine a Eomaia al discendente elefante sia in quello che va dall'antenato affine a Eomaia al discendente scimpanzé. Ma non si tratta affatto di un elefanzé. Se lo fosse, dovrebbe essere anche un canantino, perché chiunque sia l'antenato comune di uno scimpanzé e un elefante, è anche l'antenato comune di un cane e un lamantino. E dovrebbe essere pure un oritteropotamo, perché il medesimo antenato è anche l'antenato comune di un orittèropo e un ippopotamo. L'idea stessa di un canantino (o di un elefanzé, di un oritteropotarno, di un canguroceronte odi un bufaleone) è assurda e profondamente antievolutiva. Altrettanto lo è la ranimmia, ed è una vergogna che chi ha tirato fuori questa idiozia, il predicatore itinerante australiano John Mackay, abbia fatto il giro delle scuole britanniche nel 2008 (e nel 2009) spacciandosi per «geologo» e insegnando a bambini innocenti che se l'evoluzione fosse vera la documentazione fossile comprenderebbe le «ranimmie».
Un esempio altrettanto assurdo è il libro dell'apologeta dell'Islam Harun Yahya, Atlas of Creation, un tomo enorme, pubblicato senza lesinare sui costi, con illustrazioni patinate e una notevole dose di superficialità e ignoranza. Il libro è chiaramente costato una fortuna, sicché stupisce ancora di più che sia stato distribuito gratis a decine di migliaia di insegnanti di scienze, tra cui me. Nonostante tutti i quattrini che vi sono stati profusi, contiene una serie di errori clamorosi. Per esporre un concetto falso, ossia che i fossili più antichi sono indistinguibili dai loro omologhi moderni, Yahya mostra un serpente di mare definendolo «anguilla» (due animali così diversi da trovarsi addirittura in differenti classi di vertebrati), una stella di mare (asteroidei) chiamandola «stella serpentina» (nome comune di un'altra classe di echinodermi, gli ofiuroidei), un sabellide (anellidi) spacciandolo per crinoide (sabellidi e crinoidi non solo appartengono a phyla diversi, ma a sottoregni diversi, sicché non potrebbero essere più lontani, anche se sono entrambi animali) e, svarione più divertente di tutti, un'esca artificiale denominandola «friganea».
Ma oltre a queste perle di stupidità tendenziosa, il libro contiene una sezione sugli anelli mancanti. Al lettore viene sottoposta in tutta serietà un'immagine che dovrebbe dimostrare come non esistano stadi intermedi tra un pesce e una stella di mare. Non posso credere che Yahya pensi seriamente che gli evoluzionisti si aspettino di trovare forme transizionali tra due animali così diversi come una stella di mare e un pesce. Sospetto quindi che egli conosca fin troppo bene i suoi lettori e sfrutti con deliberato cinismo la loro ignoranza.
Ancora una volta ripeterò che gli esseri umani non discendono dalle scimmie. Abbiamo semplicemente un antenato in comune con le scimmie. L'antenato comune doveva somigliare molto più a una scimmia che a un uomo, e forse lo avremmo effettivamente chiamato scimmia se lo avessimo conosciuto, circa 25 milioni di anni fa. Ma per quanto l'uomo si sia evoluto da un antenato che potremmo ragionevolmente definire una scimmia, nessun animale dà origine all'improvviso a una nuova specie, o almeno a una specie diversa da lui quanto l'uomo è diverso da una scimmia non antropomorfa o anche da uno scimpanzé. Non è questo che si intende con evoluzione. L'evoluzione non solo è, di fatto, un processo graduale, ma deve esserlo se vuole svolgere un ruolo esplicativo. Grandi balzi compiuti in un'unica generazione, come quello dell'ipotetica scimmia che partorisce il bambino, sono improbabili quasi quanto la creazione divina e sono esclusi per la stessa ragione: troppo improbabili sotto il profilo statistico. Come sarebbe bello se i critici dell'evoluzione si disturbassero ad apprendere anche solo i minimi rudimenti della materia che criticano.
Alla base di gran parte delle pretestuose richieste di «anelli mancanti» vi è un mito medievale che pervase la mente degli uomini fino all'era di Darwin e ha pervicacemente continuato a confonderla anche dopo. E’ il mito della Grande catena dell'essere, secondo il quale tutto, nell'universo, è disposto a scala: in cima c'è Dio, poi vengono gli arcangeli, le varie schiere di angeli, gli uomini, gli animali, le piante, e infine le pietre e altri oggetti inanimati. Considerato che questa concezione risale a un'epoca in cui il razzismo era connaturato nell'uomo, non occorre aggiunga che gli esseri umani non occupavano tutti lo stesso piolo. No di certo. Naturalmente i maschi erano di un intero gradino sopra le femmine della loro stessa specie (ecco perché poc'anzi ho detto «pervase la mente degli uomini»). Ma fu la presunta gerarchia all'interno del regno animale a confondere di più le idee quando irruppe in scena il concetto di evoluzione. Pareva naturale supporre che gli animali «inferiori» si evolvessero in animali «superiori». E di conseguenza era logico aspettarsi di vedere «nessi» tra di loro lungo tutta l'estensione della «scala>'. Una scala con molti pioli mancanti convince poco. Ed ecco che l'immagine di una scala senza pioli diventa il leitmotiv di gran parte delle critiche sul tema degli «anelli mancanti". Ma il mito della scala è, come vedremo tra poco, del tutto fuorviante e antievolutivo.
Espressioni come «animali superiori» e «animali inferiori» ci escono di bocca così facilmente che rimaniamo sbigottiti quando ci rendiamo conto che, lungi dall'adattarsi bene, come potremmo supporre, al pensiero evolutivo, gli sono, e gli sono sempre state, profondamente antitetiche. Siamo convinti che gli scimpanzé siano animali superiori e i lombrichi animali inferiori, che il significato di questa differenza ci sia sempre stato noto e che l'evoluzione lo renda ancora più chiaro. Ma non è così. Non è affatto chiaro se questa distinzione abbia un significato; e, se ce l'ha, ne ha talmente tanti da riuscire fuorviante o addirittura perniciosa.
Ecco un elenco dei messaggi più o meno disorientanti che potremmo voler inviare quando definiamo per esempio una scimmia «superiore» a un lombrico.
1. «Le scimmie si sono evolute dai lombrichi.» E’ falso, come è falso che l'uomo si sia evoluto dagli scimpanzé. Le scimmie e i lombrichi hanno un antenato comune.
2. «L'antenato comune delle scimmie e dei lombrichi somigliava più a un lombrico che a una scimmia.» Questo messaggio ha potenzialmente più senso. Si può addirittura usare il termine «primitivo» quasi alla lettera, se con esso si intende «somigliante agli antenati», ed è naturalmente vero che alcuni animali moderni sono, in questo senso, più primitivi di altri. Se ci pensiamo bene, la frase significa che la più primitiva di due specie è cambiata meno rispetto all'antenato comune (tutte le specie, nessuna esclusa, condividono un antenato comune se si risale abbastanza indietro nel tempo). Se nessuna delle due specie è cambiata molto più dell'altra, non si dovrebbe ricorrere al termine «primitivo» quando le si confronta.
Vale la pena fermarsi un attimo a riflettere su un concetto connesso a tale conclusione. E difficile misurare i gradi di somiglianza; e in ogni caso non c'è nessuna ragione necessaria per cui l'antenato comune di due animali moderni debba somigliare più all'uno che all'altro. Dati due animali, per esempio un'aringa e un calamaro, è possibile che uno assomigli all'antenato comune più dell'altro, ma non ne consegue che debba sempre essere così. Essi hanno avuto un arco di tempo esattamente uguale per divergere dal progenitore, sicché la principale previsione di un evoluzionista sarebbe semmai che nessun animale moderno fosse più primitivo di un altro.
Potremmo aspettarci che entrambi fossero cambiati nella stessa misura, ma procedendo in direzioni diverse a partire dall'epoca dell'antenato comune. Di fatto questa previsione è spesso smentita (come nel caso della scimmia e del lombrico), ma non vi è nessun fondato motivo per prevedere che lo sia. Inoltre, le parti da cui gli animali sono composti non evolvono tutte allo stesso ritmo: un animale può essere primitivo dalla cintola in giù, ma molto evoluto dalla cintola in su. Scherzi a parte, un organismo potrebbe avere il sistema nervoso più primitivo rispetto all'altro, e l'altro avere più primitivo lo scheletro. Si noti soprattutto che «primitivo», nell'accezione di «somigliante all'antenato», non significa necessariamente «semplice» (nel senso di meno complesso). Lo zoccolo di un cavallo è più semplice del piede umano (ha un unico dito invece di cinque, tanto per citare una caratteristica), ma il piede umano è più primitivo (l'antenato che condividiamo con il cavallo aveva, come noi, cinque dita, sicché il cavallo è cambiato di più). Questo ci conduce alla voce successiva della nostra lista.
3. «Le scimmie sono più intelligenti [o più belle, o dotate di genoma più grande o piano corporeo più complesso ecc.] dei lombrichi.» Questo snobismo zoologico produce danni quando si comincia ad applicarlo scientificamente. Lo menziono solo perché viene spesso confuso con gli altri significati e il modo migliore di eliminare la confusione è denunciarla. Si possono immaginare molte scale in base alle quali classificare gli animali, non solo le quattro che ho menzionato. Gli animali che si trovano in alto sull'una potrebbero (o no) trovarsi in basso nell'altra. I mammiferi hanno sicuramente il cervello più grande delle salamandre, ma hanno il genoma più piccolo di alcune di esse.
4. «Le scimmie somigliano all'uomo più dei lombrichi.» Nel caso particolare delle scimmie e dei lombrichi, è innegabile. Ma con questo? Perché dovremmo scegliere Homo sapiens come parametro in base al quale giudicare gli altri organismi? Una sanguisuga potrebbe replicare indignata che i lombrichi hanno la grande virtù di somigliare più degli uomini alle sanguisughe. Benché tradizionalmente la Grande catena dell'essere collochi l'uomo tra l'animale e l'angelo, nulla giustifica, sotto il profilo evolutivo, la diffusa idea che l'evoluzione sia in qualche modo «mirata» all'essere umano o l'essere umano sia «l'ultima parola dell'evoluzione». E’ incredibile vedere fin dove si spinga questo assunto vanaglorioso. Al livello più rozzo esso trapela dalla querula domanda che si sente fare tante volte: «Se lo scimpanzé si è evoluto in uomo, come mai ci sono ancora in giro scimpanzé?». Vi ho già accennato e purtroppo non è uno scherzo. Mi sento rivolgere questa domanda infinite volte, in certe occasioni perfino da persone che sembrano colte.
5. «Le scimmie [e altri animali «superiori»] sono più abili a sopravvivere dei lombrichi [e altri animali «inferiori»].» Questo non è neanche lontanamente sensato, e nemmeno vero. Tutte le specie viventi sono sopravvissute almeno fino al presente. Alcune scimmie, come i mirabili tamarini dorati, corrono il rischio di estinguersi; sono quindi molto meno abili a sopravvivere dei lombrichi. I ratti e gli scarafaggi prosperano nonostante siano considerati da molti «inferiori» ai gorilla e agli orangutan, i quali sono pericolosamente vicini all'estinzione.
Spero di avere spiegato a sufficienza che è assurdo classificare le specie moderne in base a un'ipotetica scala gerarchica, come se fosse evidente quali siano le «superiori» e quali le «inferiori», e spero di avere dimostrato quanto questo atteggiamento sia antievolutivo. Si possono immaginare innumerevoli scale; a volte può anche avere senso collocare separatamente gli animali su alcune di esse, ma le scale non sono ben correlate tra loro e nessuna ha il diritto di essere definita «scala evolutiva». Abbiamo visto la vecchia tentazione a commettere errori grossolani come chiedersi: «Perché non ci sono ranimmie?». Ma il pernicioso retaggio della Grande catena dell'essere alimenta anche domande provocatorie come «Dove sono gli stadi intermedi tra i principali gruppi animali?», ed è quasi altrettanto disdicevole la tendenza di alcuni evoluzionisti a rispondere a tale domanda tirando fuori fossili come l'Archaeopteryx, il famoso «intermedio tra rettili e uccelli». Siccome però dietro la risposta fallace dell'Archaeopteryx vi è qualcos'altro, che è di importanza generale, dedicherò all'argomento ancora qualche considerazione, usando l'Archaeopteryx come esempio di un fenomeno più ampio.
Gli zoologi hanno sempre diviso tradizionalmente i vertebrati in classi, ampie categorie con nomi come mammiferi, uccelli, rettili e anfibi. Alcuni zoologi, chiamati «cladisti» [* Dal termine «clade», che significa un gruppo di organismi i cui membri discendono tutti, evolutivamente, da un antenato comune], sostengono che una classe, per essere tale, deve consistere di animali che condividono tutti un antenato comune appartenente a quella stessa classe e privo di discendenti in altra classe. Gli uccelli costituiscono un buon esempio di classe. Tutti discendono da un unico antenato che a sua volta sarebbe stato chiamato uccello da un ipotetico zoologo dell'epoca e avrebbe condiviso con gli uccelli moderni i caratteri diagnostici fondamentali: penne, ali, becco ecc. Gli animali comunemente chiamati rettili non sono, in questo senso, una buona classe, perché, almeno nella tassonomia tradizionale, la categoria esclude esplicitamente gli uccelli (che costituiscono una classe a sé) e tuttavia alcuni «rettili» (come i coccodrilli e i dinosauri) sono sempre stati ritenuti cugini più stretti degli uccelli che di altri «rettili» (come le lucertole e le tartarughe). Anzi, alcuni dinosauri sono cugini più stretti degli uccelli che di altri dinosauri. I «rettili», quindi, sono una classe artificiale, perché gli uccelli ne sono esclusi artificialmente. A rigor di termini, se volessimo fare dei rettili una classe realmente naturale, dovremmo includervi gli uccelli come rettili. Gli zoologi con tendenze cladistiche evitano del tutto il termine «rettili» e dividono questi animali in Archosauria (dinosauri, coccodrilli e uccelli), Lepidosauria (serpenti, lucertole e il raro sfenodonte della Nuova Zelanda) e Testudinidae (tartarughe marine e terrestri). Gli zoologi con tendenze non cladistiche usano volentieri termini come «rettile», perché lo trovano descrittivamente utile, anche se esclude in maniera artificiale gli uccelli.
Ma che cosa c'è, negli uccelli, che ci induce a separarli dai rettili? Perché mai conferiamo loro il nobile appellativo di «classe», quando, dal punto di vista evolutivo, sono solo un ramo della categoria rettili? Lo facciamo perché i loro cugini più stretti nell'albero della vita, i rettili a essi contigui, sono estinti, mentre gli uccelli, unici del loro genere, hanno continuato a vivere. I loro parenti più stretti si rinvengono tutti tra i dinosauri da tempo estinti. Se fosse sopravvissuta un'ampia varietà di linee ancestrali di dinosauri, gli uccelli non avrebbero attirato la nostra attenzione; non li avremmo elevati al rango di classe di vertebrati a sé stante, né avremmo fatto domande come: «Dove sono gli anelli mancanti tra rettili e uccelli?». L'Archaeopteryx sarebbe ancora un bel fossile da museo, ma non avrebbe l'odierno, cruciale ruolo di risposta di rito alla frase «Mostrateci gli stadi intermedi», che risulterebbe con grande chiarezza una vuota provocazione. Se le carte dell'estinzione fossero state distribuite in maniera diversa, vedremmo intorno a noi innumerevoli dinosauri, alcuni dei quali pennuti, volatori e dotati di becco, denominati uccelli. In effetti, si stanno scoprendo sempre più fossili di dinosauri pennuti e sta diventando molto chiaro che non esiste alcuna grande sfida degli «anelli mancanti» di cui l'Archaeopteryx rappresenti la risposta.
Occupiamoci adesso di alcune delle maggiori transizioni evolutive, i cui «anelli» si è preteso fossero «mancanti».
A parte il viaggio nello spazio a bordo di un razzo, non c'è passaggio più ardito o cambiamento di vita più radicale di quello che induce una creatura ad abbandonare il mare per la terraferma. Le due zone biotiche sono diverse sotto talmente tanti profili che spostarsi dall'una all'altra richiede un mutamento radicale di quasi tutte le parti del corpo. Branchie che servono a ricavare ossigeno dall'acqua sono inutili in aria, mentre i polmoni sono inutili nell'acqua. I metodi di propulsione che sono rapidi, eleganti ed efficienti in mare sono pericolosamente goffi sulla terraferma, e viceversa. Non c'è da stupirsi se sono divenute proverbiali espressioni quali «sentirsi come un pesce fuor d'acqua» o «nuotare come un ferro da stiro». Né c'è da stupirsi se gli «anelli mancanti» in questa regione della documentazione fossile siano di straordinario interesse.
Se si risale abbastanza indietro nel tempo, si scopre che tutte le creature vivevano in mare, l'alma mater acquea e salata di tutta la vita. In diversi momenti della storia evolutiva, individui intraprendenti di molti gruppi animali si spostarono sulla terraferma, fino ai deserti più aridi, portandosi dietro il loro mare privato sotto forma di sangue e liquidi cellulari. Oltre ai rettili, agli uccelli, ai mammiferi e agli insetti che vediamo intorno a noi, altri gruppi che sono riusciti a compiere il grande passaggio dall'utero acqueo della vita alla terraferma sono gli scorpioni, le lumache, crostacei come gli onischi e i granchi, i millepiedi e i centopiedi, i ragni e i loro affini, e almeno tre phyla di vermi. Né vanno dimenticate le piante, costanti generatrici di carbonio fruibile, senza la cui precedente invasione della terraferma non sarebbe stata possibile nessun'altra migrazione.
Per fortuna, degli stadi transizionali dell'esodo dei pesci verso la terraferma resta una bella documentazione nei fossili. Altrettanto documentati sono gli stadi transizionali che vanno nella direzione opposta e che videro gli antenati delle balene e dei dugonghi abbandonare la terraferma faticosamente conquistata per tornare nei mari ancestrali. In entrambi i casi, gli anelli che un tempo mancavano adesso abbondano e abbelliscono i nostri musei.
Quando diciamo che i «pesci» emersero sulla terraferma, dobbiamo ricordare che i «pesci», come i «rettili», non costituiscono un gruppo naturale. I pesci sono definiti per esclusione. Sono tutti i vertebrati eccetto quelli che si sono trasferiti sulla terraferma. Poiché l'intera evoluzione iniziale dei vertebrati ebbe luogo nell'acqua, non c'è da stupirsi che la maggior parte dei rami sopravvissuti dell'albero dei vertebrati si trovi ancora in mare. E li chiamiamo tuttora «pesci» anche quando sono imparentati solo alla lontana con altri «pesci». La trota e il tonno sono cugini più stretti dell'uomo che dello squalo, eppure li chiamiamo «pesci». I dipnoi e i celacanti sono più strettamente imparentati con noi che con la trota e il tonno (e naturalmente lo squalo), ma li chiamiamo sempre «pesci». Perfino gli squali sono cugini più stretti dell'uomo che delle lamprede e dei missinoidi (gli unici superstiti del gruppo un tempo fiorente e variegato dei pesci senza mascelle), ma anch'essi li chiamiamo pesci. I vertebrati i cui antenati non si avventurarono mai sulla terraferma hanno tutti l'aspetto di «pesci», nuotano tutti come pesci (diversamente dai delfini, che flettono la spina dorsale in senso verticale anziché in senso orizzontale come i pesci) e tutti, immagino, hanno sapore di pesce.
Come abbiamo appena visto nell'esempio dei rettili e degli uccelli, per un evoluzionista un gruppo «naturale» di animali è un gruppo i cui membri sono tutti più strettamente imparentati tra loro che con i non appartenenti al gruppo. Gli «uccelli» sono un gruppo naturale il cui antenato comune più recente è condiviso da tutti, ma non dai non‑uccelli. Secondo questa definizione, i «pesci» e i «rettili» non sono gruppi naturali. L'antenato comune più recente di tutti i «pesci» è condiviso anche da molte creature che non sono pesci. Se escludiamo i nostri lontani cugini squali, noi mammiferi apparteniamo a un gruppo naturale che comprende tutti i moderni pesci ossei (i quali si contrappongono ai pesci cartilaginei, come gli squali). Se poi mettiamo da parte i pesci ossei attinopterigi (salmone, trota, tonno, pesce angelo: quasi tutti i pesci che vediamo in mare, a parte gli squali), il gruppo naturale a cui apparteniamo comprende tutti i vertebrati di terra più i cosiddetti crossopterigi. E proprio dalle file dei crossopterigi che proveniamo noi, ed è venuto il momento di dedicare loro particolare attenzione.
I crossopterigi, oggi, comprendono solo i dipnoi e i celacanti (si sono «ridotti» in quanto «pesci», ma sulla terraferma si sono assai espansi: noi vertebrati di terra siamo dipnoi aberranti). Sono «crossopterigi» perché dotati di pinne che, anziché avere lo scheletro a raggio dei pesci a noi più noti, sono come zampe. Non a caso Old Fourlegs (Antichi quadrupedi) era il titolo di un libro popolare sui celacanti scritto da J.L.B. Smith, il biologo sudafricano che più di tutti gli altri attirò l'attenzione del mondo su questi animali dopo che, nel 1938, venne incredibilmente scoperto il primo celacanto vivo nelle reti di un peschereccio sudafricano. «Ero non meno stupito che se avessi visto camminare per la strada un dinosauro» disse. Si conoscevano già celacanti fossili, ma li si riteneva estinti fin dall'epoca dei dinosauri. Smith descrisse con parole commoventi il momento in cui, convocato per una consulenza da Margaref Latimer, scopritrice del reperto vivente (in suo onore chiamò poi l'animale Latimeria), vide per la prima volta quel pesce straordinario:
Andammo subito al museo. La signorina Latimer era momenta neamente assente e il custode ci fece entrare nella stanza interna.
E lì c'era... un celacanto, sì, perdio! Benché fossi preparato, vederlo fu per me un'emozione fortissima: tremavo, mi girava la testa e avevo le membra informicolite. Ero assolutamente incredulo. Sì, non vi era ombra di dubbio: scaglia per scaglia, osso per osso, pinna per pinna, era un vero celacanto. Era come se una creatura di duecento milioni di anni prima fosse resuscitata. Dimentico di tutto il resto, lo rimirai, poi, quasi con timore, mi avvicinai, lo toccai e lo accarezzai, mentre mia moglie mi guardava in silenzio. Arrivò la signorina Latimer e mi salutò cordialmente. Solo allora mi tornò la favella; non ricordo bene che cosa dissi, ma credo di avere spiegato ai presenti che era autentico, assolutamente autentico, un indiscusso celacanto. Non potevo più dubitarne.
I celacanti sono più strettamente imparentati con noi che con la maggior parte dei pesci. Sono un po' cambiati dall'epoca del nostro antenato comune, ma non abbastanza da essere esclusi dalla categoria degli animali che nel linguaggio colloquiale e in quello dei pescatori sono definiti «pesci». Tuttavia, assieme ai dipnoi, sono cugini assai più stretti dell'uomo che di trote, salmoni, tonni e la maggior parte dei pesci. Celacanti e dipnoi sono esempi di «fossili viventi».
Beninteso, non discendiamo dai dipnoi o dai celacanti. Condividiamo con i dipnoi un antenato che somigliava più a loro che all'uomo, ma che non ricordava molto né gli uni né l'altro. I dipnoi saranno anche fossili viventi, ma non assomigliano molto ai nostri antenati. Nella ricerca dei nostri progenitori, dobbiamo invece orientarci verso i fossili racchiusi nelle rocce. In particolare, ci conviene interessarci ai fossili del Devoniano che radiografano la transizione tra i pesci rimasti in ambiente acquatico e i primi vertebrati approdati sulla terraferma. Peccheremmo di eccessivo ottimismo se pensassimo di trovare tra quei fossili i nostri antenati, ma possiamo sperare di rinvenire loro cugini abbastanza stretti da suggerirci che aspetto avessero.
Una delle lacune più famose della documentazione fossile, talmente cospicua da aver ricevuto un nome, «lacuna di Romer» (dal nome del celebre paleontologo americano A.S. Romer), riguarda un periodo che va da 360 milioni di anni fa, alla fine del Devoniano, a 340 milioni di anni fa, all'inizio del Carbonifero. Dopo la lacuna di Romer, troviamo indubbi anfibi che strisciano per le paludi, una ricca irradiazione di animali simili a salamandre, che in alcuni casi erano grandi come coccodrilli, ai quali assomigliavano superficialmente. Pare sia stata un'epoca di giganti, perché c'erano libellule con un'apertura alare pari all'estensione del mio braccio: gli insetti più grandi che siano mai vissuti.* [ Si è formulata l'ipotesi che il gigantismo sia stato reso possibile dal maggiore contenuto di ossigeno nell'atmosfera dell'epoca. Gli insetti non hanno polmoni e respirano per mezzo di tubuli che pompano l'aria nel corpo. I tubuli dell'aria non possono costituire un sistema di distribuzione così esteso e complesso come quello dei vasi sanguigni, ed è possibile che questo limiti le dimensioni corporee. Le dimensioni sarebbero state più grandi se l'atmosfera avesse contenuto ii 35% di ossigeno, anziché il 21% di oggi. L'alta percentuale di ossigeno giustificherebbe le libellule giganti, ma non è detto che sia la spiegazione giusta. Per inciso, mi stupisce che, con tutto quell'ossigeno in giro, non siano scoppiati continui incendi. Forse scoppiavano. Gli incendi delle foreste dovevano essere assai più frequenti di oggi e i fossili rivelano un'alta percentuale di specie vegetali resistenti al fuoco. Non si sa bene perché il contenuto d'ossigeno dell'atmosfera abbia raggiunto un picco durante il Carbonifero e il Permiano;forse perché è stato sequestrato tanto carbonio sotto forma di carbone del sottosuolo.]
A cominciare da 340 milioni di anni fa, potremmo quasi chiamare il Carbonifero l'equivalente anfibio dell'era dei dinosauri. Prima di allora, però, vi è la lacuna di Romer; e prima di quella, Romer vedeva solo pesci, crossopterigi che vivevano nell'acqua. Dov'erano gli intermedi e che cosa indusse i crossopterigi ad avventurarsi sulla terraferma?
La mia fantasia di studente, a Oxford, si accendeva quando andavo alle lezioni dell'eruditissimo Harold Pusey, il quale, nonostante l'oratoria arida e prolissa, aveva il dono di vedere, al di là delle nude ossa, gli animali di carne e sangue che erano vissuti in un mondo passato. Quando spiegò che cosa poteva aver indotto alcuni crossopterigi a sviluppare polmoni e zampe, facendo sua un'ipotesi dello stesso Romer, la sua lezione parve illuminante alle mie orecchie di allievo e ancora la condivido, anche se l'idea è meno in auge presso i paleontologi moderni di quanto non lo fosse all'epoca di Romer. Romer e Pusey immaginarono siccità annuali durante le quali laghi, stagni e torrenti si prosciugavano, per poi gonfiarsi di nuovo l'anno seguente. Alcuni pesci, fra i tanti che si trascinavano da un lago o da uno stagno che si stavano prosciugando verso acque più profonde in cui poter sopravvivere fino alla successiva stagione umida, acquisirono una temporanea abilità a resistere in ambiente terrestre. Secondo tale ipotesi, quello dei nostri antenati non fu tanto un trasferimento sulla terraferma, quanto un uso della terraferma come ponte temporaneo per tuffarsi di nuovo in acqua. Molti animali moderni fanno lo stesso.
Purtroppo, però, Romer aveva esposto la sua teoria insieme alla premessa che il Devoniano era un'epoca di siccità. Così, quando prove successive dimostrarono che la premessa era errata, sembrò errata l'intera teoria. Romer avrebbe fatto meglio a rinunciare a quella ridondante premessa. Come ho osservato nel Racconto dell'antenato, la teoria funziona ancora, anche se il Devoniano è risultato meno funestato da siccità di quanto Romer pensasse.
Torniamo ai fossili. Sono presenti in forma sparsa per tutto il tardo Devoniano, il periodo immediatamente precedente il Carbonifero: meravigliose tracce di «anelli mancanti», animali che cominciarono a colmare il divario tra i crossopterigi, così abbondanti nei mari devoniani, e gli anfibi che in seguito avrebbero strisciato nelle paludi del Carbonifero. Sul versante ittico del divario, nel 1881 fu scoperto l'Eusthenopteron (fig. 6.3) in una collezione di fossili provenienti dal Canada.
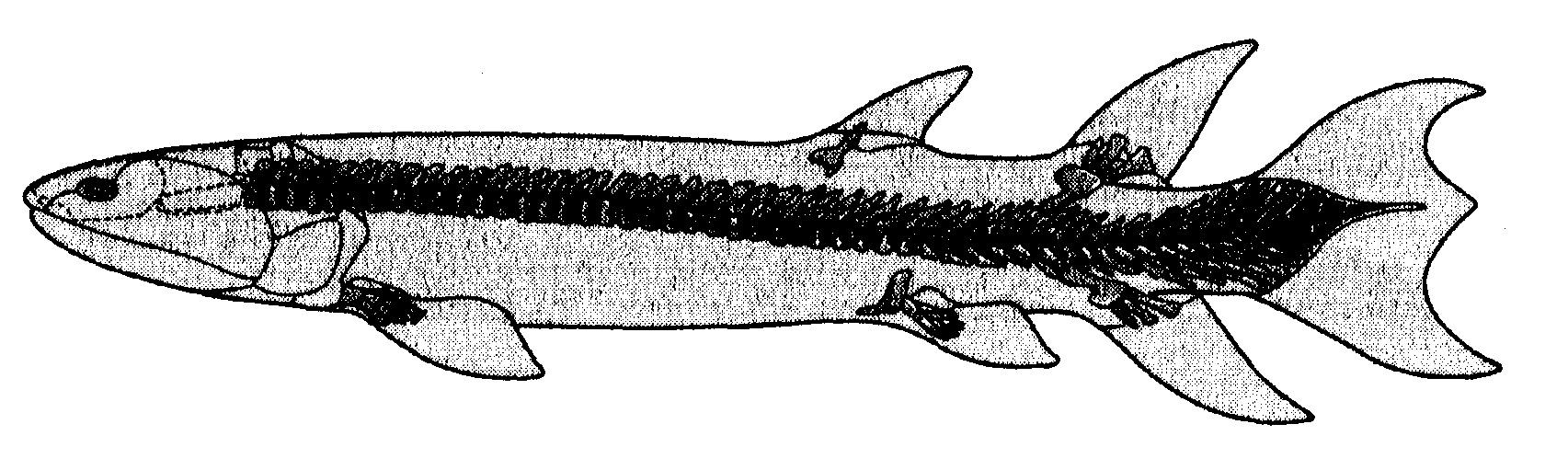
Pare sia stato un pesce che cacciava in superficie e forse, nonostante alcune fantasiose ricostruzioni ottocentesche, non approdò mai sulla terraferma. Nondimeno presentava diverse somiglianze anatomiche con gli anfibi di 50 milioni di anni dopo, tra cui le ossa del cranio, i denti e, soprattutto, le pinne. Benché fossero usate con tutta probabilità per nuotare e non per camminare, le ossa avevano la tipica struttura di quelle dei tetrapodi (il nome dato a tutti i vertebrati di terra). Nelle pinne pettorali, un unico omero era unito a due ossa, il radio e l'ulna, collegati a molti ossicini che noi tetrapodi chiameremmo ossa carpali, metacarpali e dita. Inoltre, le pinne dorsali rivelavano un'analoga struttura da tetrapode.
Nel 1932 fece grande scalpore una scoperta che riguardava un'epoca vicina al versante anfibio del divario, circa venti milioni di anni dopo, al confine tra Devoniano e Carbonifero: in Groenlandia fu rinvenuto un Ichthyostega (fig. 6.4).
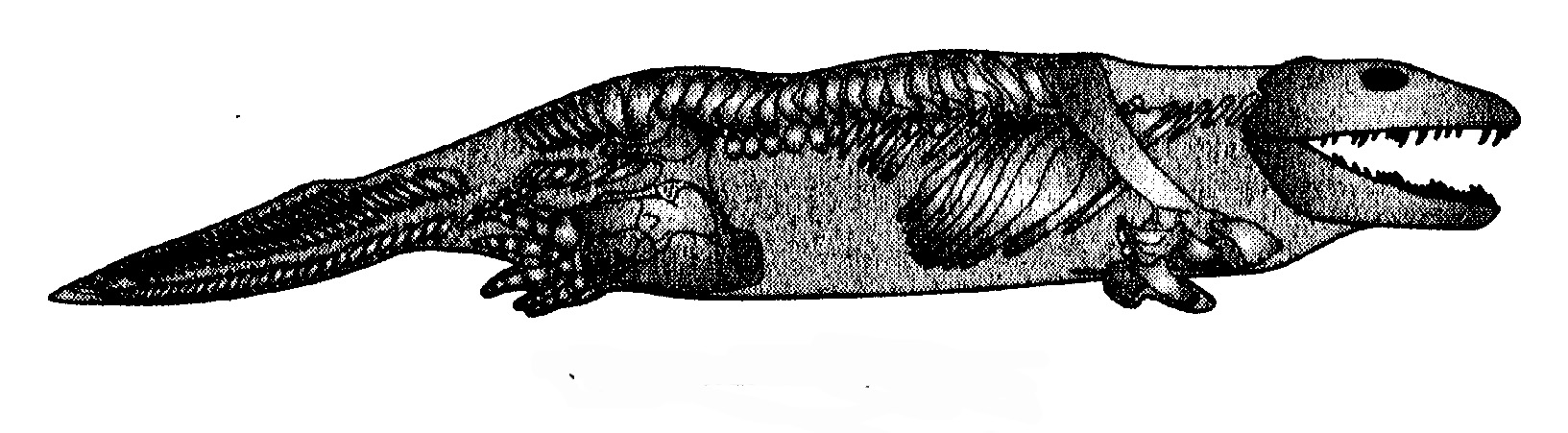
Per inciso, non si pensi a una gelida Groenlandia ricoperta di ghiacci: all'epoca dell'Ichthyostega l'isola era all'equatore. Ichthyostega fu ricostruito per la prima volta nel 1955 dal paleontologo svedese Erik Jarvik, e anche in quel caso venne rappresentato come un animale più terrestre che acquatico, il contrario di quanto fanno gli esperti odierni. La ricostruzione più recente, a opera di Per Ahlberg, anche lui, come Jarvik, dell'Università di Uppsala, pone Ichthyostega prevalentemente in acqua, anche se gli concede occasionali incursioni sulla terraferma. Tuttavia l'animale somigliava più a una salamandra gigante che a un pesce e aveva la testa piatta caratteristica degli anfibi. Diversamente da tutti i tetrapodi moderni, che hanno cinque dita dei piedi e delle mani (almeno allo stadio embrionale, anche se ne può perdere qualcuno a maturazione avvenuta), Ichthyostega aveva sette dita dei piedi. Pare che i primi tetrapodi fossero più liberi di «fare esperimenti» con un numero variabile di dita di quanto non lo siamo noi oggi. Forse a un certo punto i processi embriologici si fissarono sul numero cinque e fu compiuto un passo dal quale sarebbe stato difficile tornare indietro; anche se, bisogna ammetterlo, non così difficile: vi sono singoli gatti, e anche esseri umani, che hanno sei dita dei piedi. Le dita in più sono probabilmente dovute a un errore di duplicazione allo stadio embrionale.
Un'altra entusiasmante scoperta, anch'essa proveniente dalla Groenlandia tropicale e anch'essa risalente al confine tra Devoniano e Carbonifero, fu l'Acanthostega (fig. 6.5).
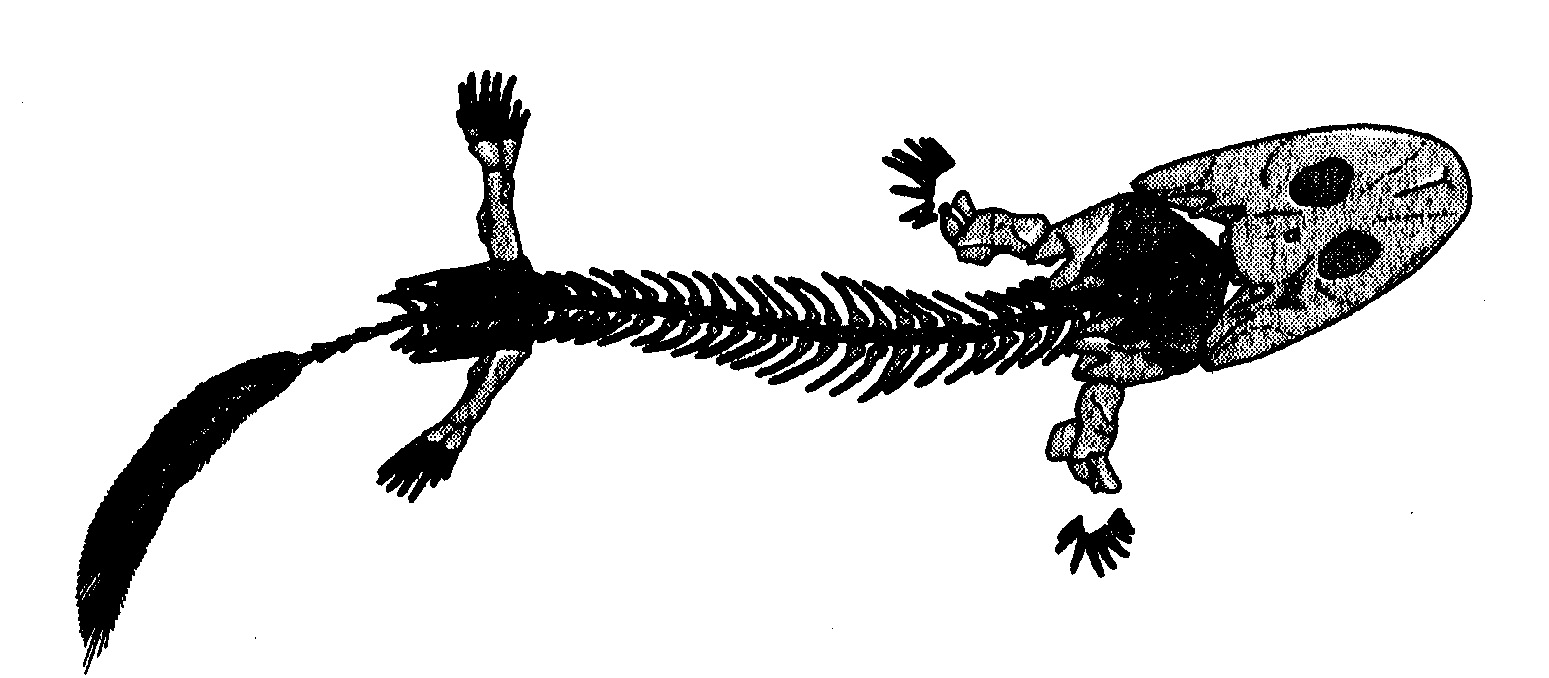
Anche Acanthostega aveva un cranio piatto da anfibio e membra da tetrapode, ma, come e più di Ichthyostega, non corrispondeva a quello che oggi reputiamo lo standard delle cinque dita, perché di dita ne aveva otto. Gli scienziati che più lo hanno studiato, Jenny Clack e Michael Coates dell'Università di Cambridge, sono convinti che, come Ichthyostega, Acanthostega fosse soprattutto un animale acquatico, ma polmoni e membra lasciano pensare che, volendo, fosse in grado di vivere sulla terraferma conio stesso perfetto agio con cui viveva in acqua. Anch'esso somigliava molto a una salamandra gigante. Spostandoci di nuovo indietro, verso il versante ittico del divario, pure Panderichthys (fig. 6.6), sempre del tardo Devoniano, è leggermente più simile a un anfibio e leggermente meno simile a un pesce di Eusthenopteron. Ma se lo vedessimo tenderemmo senz'altro a definirlo più un pesce che una salamandra.
Resta dunque una lacuna tra Panderichthys, il pesce simile a un anfibio, e Acanthostega, l'anfibio simile a un pesce. Dov'è l'«anello mancante» tra di loro? Un'équipe di scienziati dell'Università della Pennsylvania, di cui facevano parte tra l'altro Neil Shubin ed Edward Daeschier, ha deciso di scoprirlo. Shubin si è servito di questa ricerca come base per una serie di riflessioni sull'evoluzione umana nel suo libro Il pesce che è in noi. L'équipe ha riflettuto bene su quale potesse essere il luogo migliore in cui indagare e dopo una ricerca oculata ha scelto un'area rocciosa del tardo Devoniano nell'Artico canadese. Sul luogo ha trovato una miniera d'oro zoologica, il Tiktaalik,9un nome che non si dimentica, una parola inuit che significa «grande pesce d'acqua dolce». Quanto al nome specifico, roseae, mi si permetta di raccontare un aneddoto che mi riguarda. Quando sentii per la prima volta quel nome e vidi le foto (ill. lObc), pensai immediatamente al Devoniano, alla «vecchia arenaria rossa» che dà il caratteristico colore all'eponima contea del Devon e che è anche il colore di Petra («Una città rosso‑rosa, antica come metà del tempo»).
Ahimè, mi sbagliavo in pieno. La fotografia esagera la sfumatura rosata. Il nome è stato scelto in onore del benefattore che aveva contribuito a finanziare la spedizione nel Devoniano artico. Edward Daeschler è stato così gentile da mostrarmi Tiktaalik roseae quando ho pranzato con lui a Filadelfia poco dopo la sua scoperta, e lo zoologo connaturato in me, o forse il pesce che è in me, si è così commosso da rimanere senza parole. Mi pareva, forse per quel colore, di contemplare il volto del mio diretto antenato. Solo una sensazione, eppure, quel fossile, non così roseo come avevo creduto, era forse quanto di più vicino a un vero antenato estinto ‑ antico come metà del tempo ‑ mi sarebbe mai stato dato di vedere nella vita.
Se un vero Tiktaalik vivo comparisse a pochi passi da noi, forse faremmo un salto indietro spaventati come davanti a un coccodrillo, perché aveva un muso simile a quello del coccodrillo: una testa di coccodrillo su un tronco di salamandra, terminante con la parte posteriore e la coda di un pesce. Diversamente dai pesci, però, Tiktaalik aveva il collo ed era in grado di girare la testa. E’, pressoché in ogni particolare, il perfetto anello mancante: perfetto perché colma quasi con precisione il divario tra pesci e anfibi e perché non manca più. Abbiamo il fossile. Possiamo vederlo, toccano, cercare di valutarne l'età... e non riuscirci.
Il trasferimento dall'acqua alla terraferma diede luogo a una grande ridefinizione di ogni aspetto della vita, dalla respirazione alla riproduzione: fu un grande viaggio nello spazio biologico. Tuttavia, per una sorta di crudele bizzarria, parecchi perfetti animali di terra in seguito invertirono la rotta, abbandonando la riorganizzazione terrestre duramente conquistata e marciando di nuovo in acqua. Le foche e le otarie, tornate indietro solo parzialmente, ci mostrano come devono essere stati gli stadi intermedi sulla strada che porta a casi estremi come quelli delle balene e dei dugonghi. Le balene (tra cui le piccole balene che chiamiamo delfini) e i dugonghi, con i loro stretti cugini lamantini (ill. llde), abbandonarono del tutto la condizione di creature di terra per adottare di nuovo le abitudini marine dei loro remoti antenati. Oggi non tornano sulla spiaggia nemmeno per riprodursi; tuttavia, non avendo mai sviluppato nulla di equivalente alle branchie dei lontani progenitori marini, continuano a respirare aria. Altri animali che sono tornati dalla terra all'acqua o che passano in acqua parte del tempo sono le lininee, le argironete, gli scarabei d'acqua, i coccodrilli, le lontre, i serpenti di mare, i topiragni d'acqua, i cormorani non volatori delle Galapagos, le iguane marine delle Galapagos, gli opossum acquatici (marsupiali del Sudamerica), glì ornitorinchi, i pinguini e le tartarughe di mare.
Le balene sono state per lungo tempo un enigma, ma di recente abbiamo appreso molto di più sulla loro evoluzione. Prove genetiche molecolari (v. cap. X) dimostrano che i loro cugini viventi più stretti sono, in ordine decrescente, gli ippopotami, i maiali e i ruminanti. Inoltre dimostrano, con nostra grande sorpresa, che gli ippopotami sono più strettamente imparentati con le balene che con gli animali dallo zoccolo fesso (come maiali e ruminanti), fisicamente assai più simili a loro. E’ un altro esempio dell'errata associazione che a volte si fa tra grado di parentela e grado di somiglianza fisica. Abbiamo osservato il fenomeno a proposito dei pesci che risultano cugini più stretti dell'uomo che di altri pesci. In quel caso l'anomalia è sorta perché la nostra linea evolutiva ha abbandonato l'acqua per la terraferma e quindi, evolvendosi, si è allontanata sempre più dagli stretti cugini dipnoi e celacanti, i quali, rimanendo in mare, hanno continuato ad assomigliare ai nostri parenti più remoti. Ora ci ritroviamo davanti allo stesso fenomeno, ma di segno contrario. Gli ippopotami sono rimasti, almeno in parte, sulla terraferma, e così assomigliano ancora ai loro lontani cugini di terra, i ruminanti, mentre altri loro parenti più stretti, come le balene, tornando in mare sono cambiati in maniera così drastica che le loro affinità con gli ippopotami sono sfuggite a tutti i biologi eccetto i genetisti molecolari. Come avevano fatto i loro lontani progenitori pesci quando avevano preso la direzione della terraferma, così gli antenati delle balene salirono in certo modo nello spazio a bordo di un razzo o almeno di una mongolfiera quando ruppero gli ormeggi con la terraferma, liberandosi del peso vincolante della gravità.
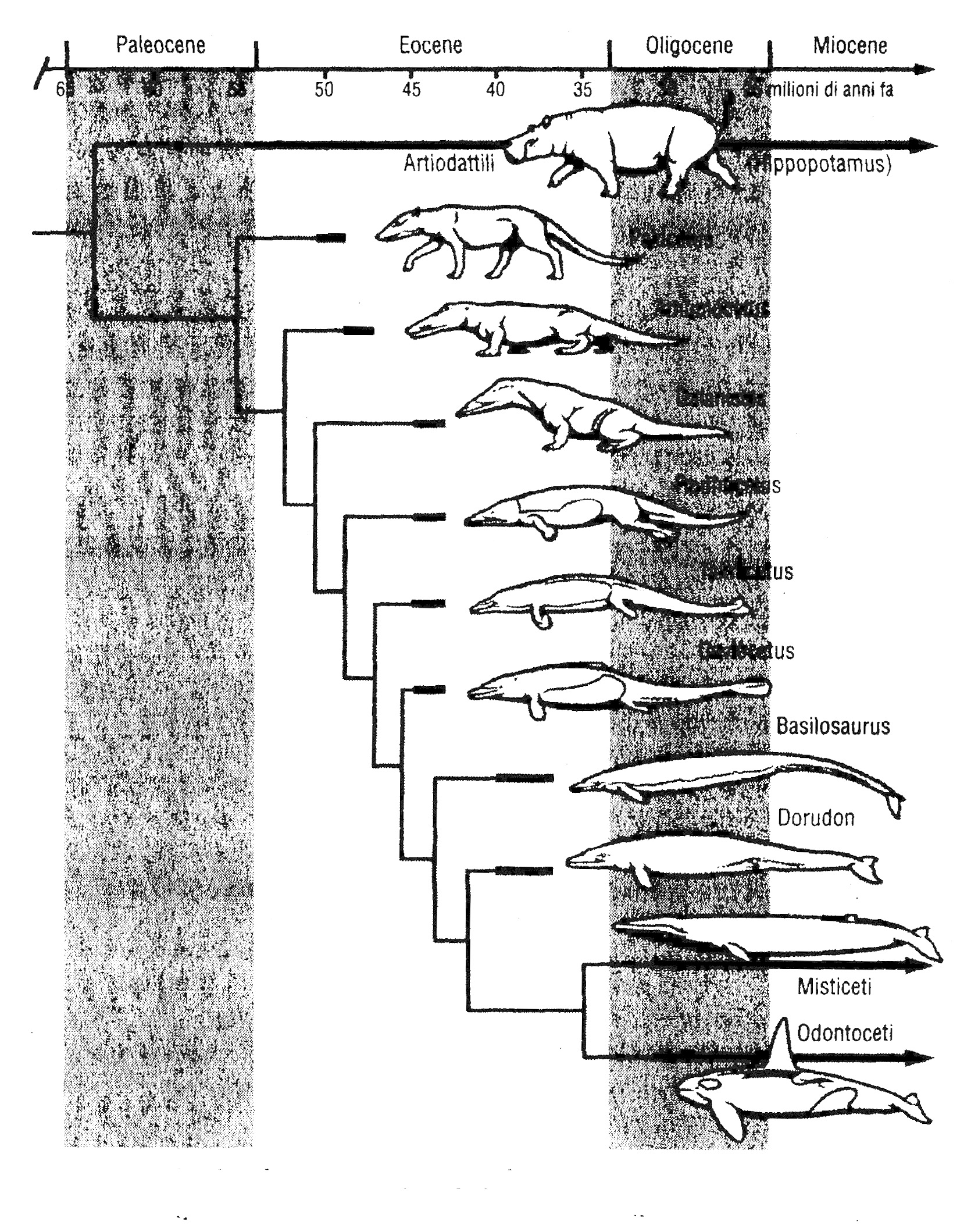
Nel contempo, la documentazione fossile sull'evoluzione della balena, in.passato piuttosto scarsa, si è arricchita di nuovi, validi tesori provenienti soprattutto dal Pakistan. La storia delle balene fossili è stata però così ben raccontata da libri come Evolution: What the
Fossils Say and Why it Matters, di Donald Prothero, e, più di recente, Why Evolution is True, di Jerry Coyne, che mi limito a suggerirne la lettura riportando soltanto un grafico (fig. 6.7) tratto dal saggio di Prothero, in cui è mostrata una sequenza di fossili in ordine cronologico. Si noti con quanta cura sono disegnati gli animali. Viene la tentazione di fare quello che facevano i libri di qualche tempo fa: tracciare frecce che vadano dai fossili più antichi ai più recenti. Ma nessuno può dire, per esempio, se Ambulocetus discenda da Pakicetus o se Basilosaurus discenda da Rod hocetus. Seguendo una politica più cauta, il grafico ipotizza invece che le balene discendano da un cugino contemporaneo di Ambulocetus il quale forse somigliava ad Ambulocetus (o era addirittura Ambulocetus). I fossili del grafico rappresentano vari stadi dell'evoluzione della balena. La graduale scomparsa degli arti posteriori, la trasformazione degli arti anteriori per la deambulazione in pinne natatorie e il progressivo appiattimento della coda fino alla trasformazione definitiva in coda di balena sono tra i cambiamenti emersi in elegante cascata.
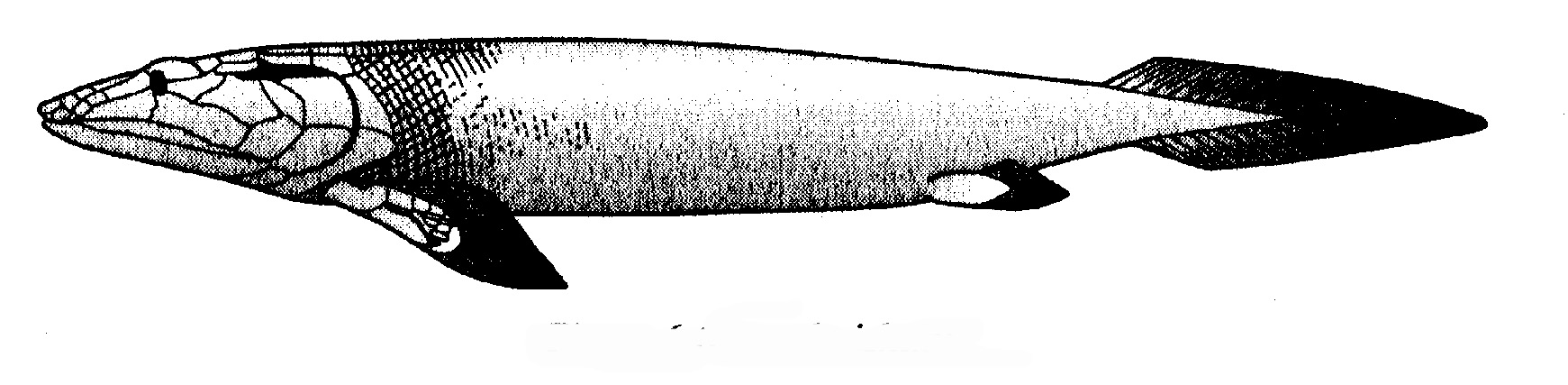
L'altro gruppo di mammiferi marini, meno numeroso e diversificato ma sempre esclusivamente acquatico, è quello dei sirenii, comprendente i dugonghi e i lamantini. Non è altrettanto ben rappresentato nella documentazione fossile, ma di recente è stato scoperto uno splendido «anello mancante». Circa contemporaneo di Ambulocetus, la «balena ambulante» dell'Eocene, è Pezosiren, il «lamantino ambulante» fossilizzatosi in Giamaica. Somiglia molto a un lamantino o a un dugongo, solo che ha quattro zampe, mentre i sirenii hanno anteriormente pinne e posteriormente nessun arto. Si vedano le figure 6.8 e 6.9.
Come le balene sono imparentate con gli ippopotami, così i sirenii sono imparentati con gli elefanti, e lo testimoniano innumerevoli prove, tra cui l'importantissima prova molecolare. Pezosiren, però, viveva forse come un ippopotamo, passando gran parte del tempo in acqua e usando le zampe sia per camminare sul fondo sia per nuotare. Il cranio era inequivocabilmente da sirenio. Non è detto che sia, come parrebbe essere, il vero antenato dei moderni lamantini e dugonghi, ma certo è incredibilmente adatto al ruolo.
Questo libro stava per andare in stampa, quando la rivista «Nature» ha dato una notizia entusiasmante: è stato scoperto nell'Artico canadese un nuovo fossile che colma una lacuna nell'ascendenza di foche, otarie e trichechi moderni (chiamati collettivamente «pinnipedi»). Completo al 65%, lo scheletro di Puijila darwini risale all'inizio del Miocene (circa 20 milioni di anni fa), un'epoca abbastanza recente perché la carta geografica del mondo fosse quasi la stessa di oggi. Dunque questa foca‑otaria primordiale (non avevano ancora cominciato a divergere) era un animale artico che viveva nelle acque fredde. E probabile che sia vissuto e abbia pescato pesci in acqua dolce (come la maggior parte delle lontre, tranne le famose lontre marine della California), anziché in mare (come la maggior parte delle foche moderne, tranne la famosa foca del Baikal). Puijila non aveva pinne natatorie, ma piedi palmati. Sulla terraferma correva probabilmente come un cane (in questo differenziandosi parecchio dai pinnipedi moderni), ma passava gran parte del tempo in acqua, dove nuotava sempre come un cane, quindi secondo uno stile assai diverso dai due adottati dalle foche e dalle otarie moderne. Puijila riempie nettamente la lacuna tra terraferma e acqua nei progenitori dei pinnipedi. E’ un'altra, deliziosa voce che si aggiunge all'elenco sempre più lungo di «anelli» non più mancanti.
Ora vorrei occuparmi di un altro gruppo che tornò dalla terraferma all'acqua: è un esempio particolarmente interessante, perché alcuni di questi animali in seguito invertirono la rotta e tornarono alla terraferma una seconda volta... Per un aspetto importante, le tartarughe marine sono meno acquatiche delle balene e dei dugonghi, perché continuano a deporre le uova sulla spiaggia. Come tutti i vertebrati ritornati in mare, le tartarughe non hanno smesso di respirare aria, ma alcune di esse sono state più abili delle balene. Ricavano infatti ulteriore ossigeno dall'acqua tramite due camere riccamente vascolarizzate poste nell'estremità posteriore del corpo. In effetti esiste una tartaruga di fiume australiana che trae la maggior parte dell'ossigeno respirando (come non esiterebbe a dire un australiano) con il culo.
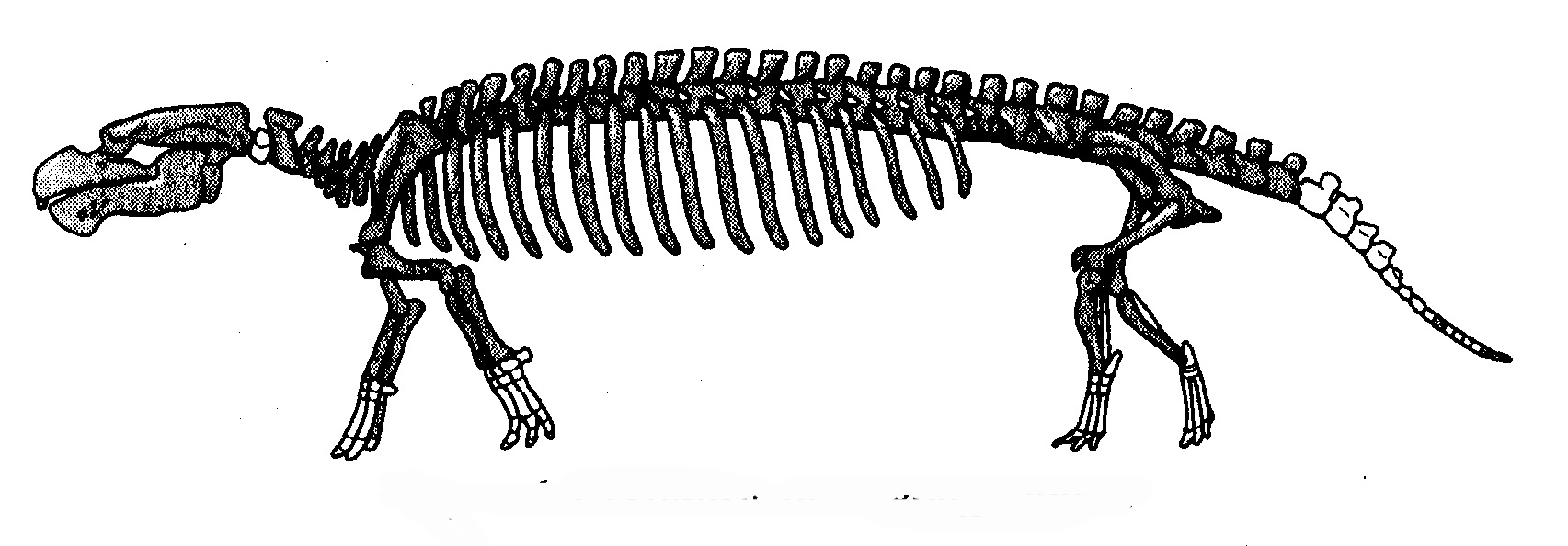
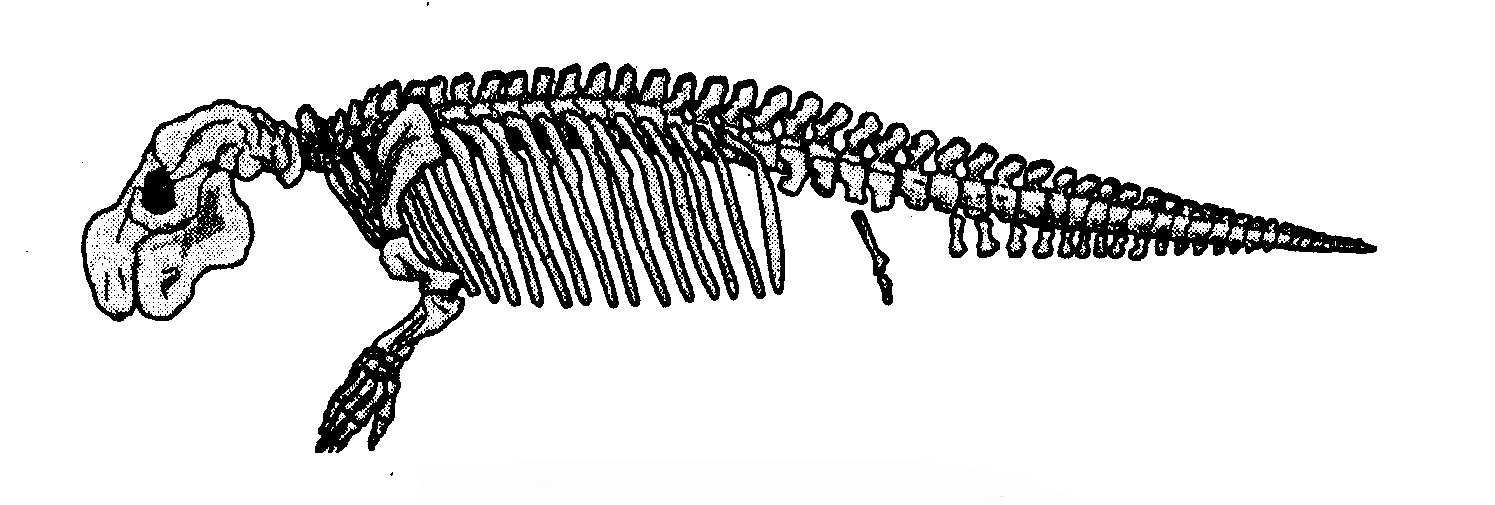
La caratteristica più singolare dei cheloni è la corazza. Come si è evoluta e che aspetto avevano gli stadi intermedi? Dove sono gli anelli mancanti? A che cosa potrebbe mai servire (domanderebbe uno zelante creazionista) mezza corazza? Be', guarda caso, è stato appena descritto sulla rivista «Nature» un nuovo fossile che risponde in maniera eloquente alla domanda. E’ una tartaruga acquatica che è stata rinvenuta negli strati del tardo Triassico in Cina e che si valuta abbia 220 milioni di anni. Si chiama Odontochelys semitestacea, dal che il lettore dedurrà come, diversamente dalla tartaruga moderna (terrestre, marina o anfibia), avesse i denti e possedesse in effetti solo mezza corazza. Aveva anche una coda molto più lunga di quella di qualsiasi chelone moderno. Tutte e tre queste caratteristiche rappresentano un eccellente materiale da «anello mancante». Il ventre era coperto da un guscio, il cosiddetto piastrone, assai simile a quello delle moderne tartarughe marine; ma mancava quasi del tutto la parte dorsale della corazza, il carapace. Il dorso era con tutta probabilità tenero come quello di una lucertola, anche se, come nel coccodrillo, vi erano alcune dure placche ossee lungo la linea mediana sopra la spina dorsale, e le vertebre erano appiattite come se «cercassero» di costituire gli inizi evolutivi di un carapace.
Accennerò a questo punto a un'interessante controversia. Gli autori dell'articolo con cui l'Odontochelys è stato presentato al mondo, Li, Wu, Rieppel, Wang e Zhao (li chiamerò per brevità gli autori cinesi, anche se Rieppel non lo è), sono convinti che Odontochelys avesse evoluto il piastrone in acqua e si stesse evolvendo verso la corazza completa. «Nature» ha la lodevole abitudine di affidare a esperti diversi dagli autori il compito di scrivere un commento sugli articoli più interessanti della settimana, che pubblica in una rubrica chiamata «News and Views». Il commento di «News and Views» sull'articolo che parlava di Odontochelys era di due biologi canadesi, Robert Reisz e Jason Head, che formulavano un'ipotesi diversa. Forse l'intera corazza si era già evoluta sulla terraferma, prima che gli antenati di Odontochelys tornassero in acqua; e forse Odontochelys aveva perso il carapace dopo essere tornato in acqua. E, dato che alcune odierne tartarughe marine, per esempio la gigantesca tartaruga liuto (Dermochelys coriacea), hanno perso o grandemente ridotto il carapace, l'ipotesi suona abbastanza plausibile.
Devo fare una breve digressione per rispondere alla domanda «A che serve mezza corazza?», ma anche a un'altra domanda: «Perché Odontochelys aveva lo scudo ventrale ma non quello dorsale?». Forse era minacciato da un pericolo proveniente dal basso, il che suggerirebbe che passasse molto tempo a nuotare vicino alla superficie, dove d'altronde doveva respirare. Gli squali oggi attaccano spesso dal basso, e non vi è motivo di supporre che all'epoca le loro abitudini di caccia fossero diverse; dunque potrebbero avere rappresentato una seria minaccia nel mondo di Odontochelys. Come esempio parallelo si può fare quello di uno dei più sorprendenti risultati dell'evoluzione, i due occhi in più del pesce Bathylychnops (fig. 6.10), che probabilmente avevano lo scopo di rilevare attacchi di predatori provenienti dal basso.
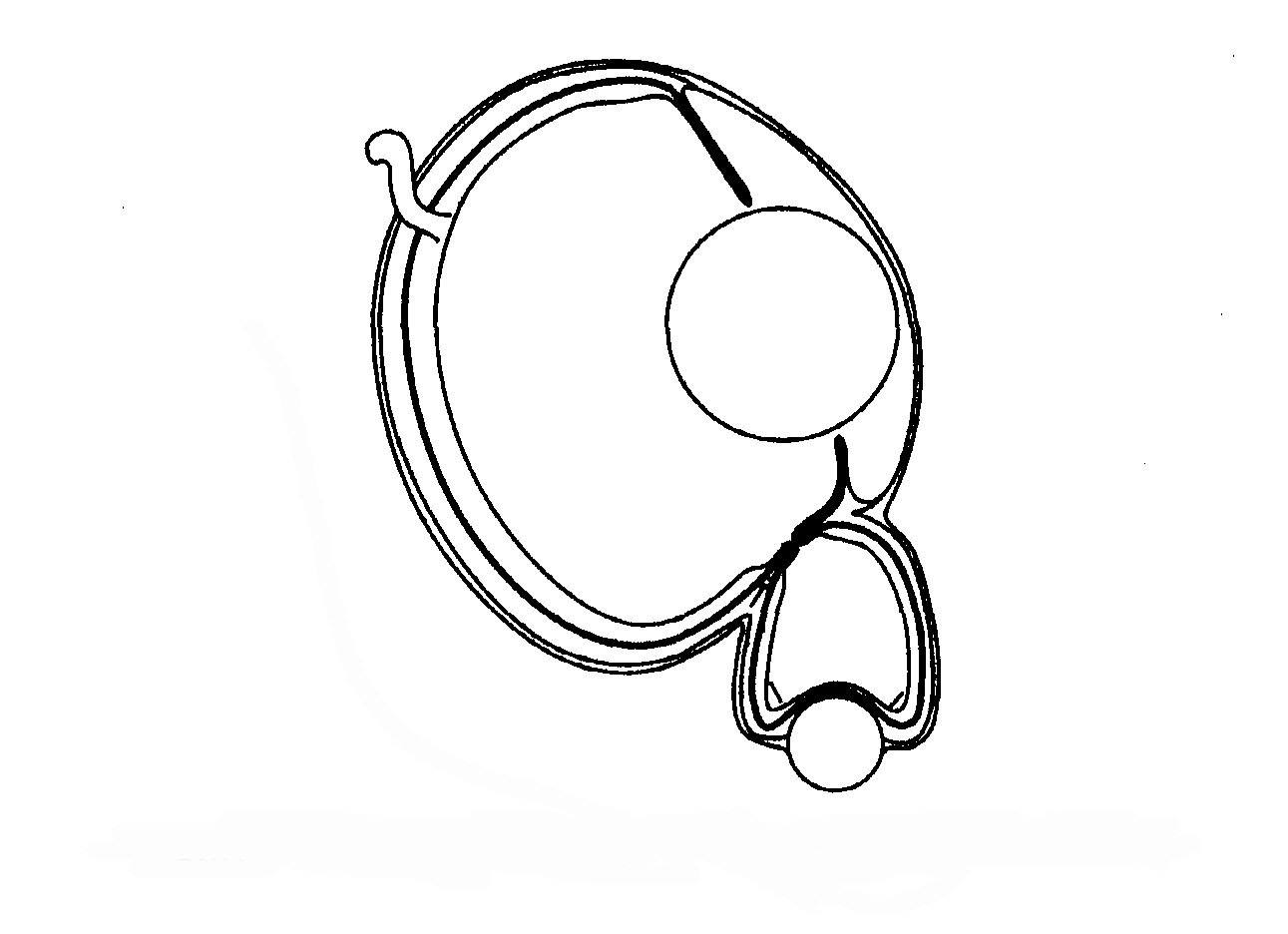
Gli occhi principali guardano di lato, come in qualsiasi pesce, ma nel lato inferiore di ciascun occhio è inserito un piccolo occhio in più, completo di cristaffino e retina, rivolto verso il basso. Se Bathylychnops si è preso il disturbo (su, non siate pedanti, avete capite che voglio dire) di sviluppare un paio di occhi in più, presumibilmente per guardarsi dagli attacchi provenienti dal basso, mi pare del tutto plausibile che per la stessa ragione Odontochelys abbia evoluto un piastrone. Il piastrone ha perfettamente senso. E se a qualcuno venisse in mente di dire «Sì, ma perché non sviluppare anche, per maggior sicurezza, uno scudo dorsale?», la risposta sarebbe facile. Gli scudi sono pesanti e ingombranti, costa svilupparli e costa portarli in giro. Ci sono sempre compromessi nell'evoluzione. Nelle tartarughe terrestri, il compromesso ha finito per favorire una corazza forte e pesante sia dorsale sia ventrale. In molte tartarughe marine, ha portato a un robusto piastrone ventrale ma a un carapace dorsale più leggero. Ed è un'ipotesi plausibile che Odontochelys abbia spinto la tendenza un poco più in là.
Se invece avessero ragione gli autori cinesi a dire che Odontochelys era in procinto di evolvere una corazza completa e che il piastrone si evolse in acqua, ne conseguirebbe che le moderne tartarughe terrestri, dotate di corazze ben sviluppate, discenderebbero dalle tartarughe marine. Come vedremo, probabilmente è vero. Ma è rilevante, in quanto significa che le odierne tartarughe terrestri rappresenterebbero una seconda migrazione dal mare alla terraferma. Nessuno ha mai sostenuto che le balene o i dugonghi siano tornati sulla terraferma dopo avere invaso il mare. L'alternativa è che le tartarughe terrestri siano sempre state sulla terraferma e abbiano evoluto la corazza in maniera indipendente, parallelamente alle loro cugine marine. Non è affatto impossibile; ma abbiamo buoni motivi per credere che le tartarughe marine siano effettivamente tornate sulla terraferma per tentare una seconda volta di diventare terrestri.
Se proviamo a tracciare l'albero filogenetico (fig. 6.11) di tutte le tartarughe marine e terrestri moderne basandoci su dati molecolari e d'altro tipo, vediamo che quasi tutti i rami sono acquatici (tipo normale).
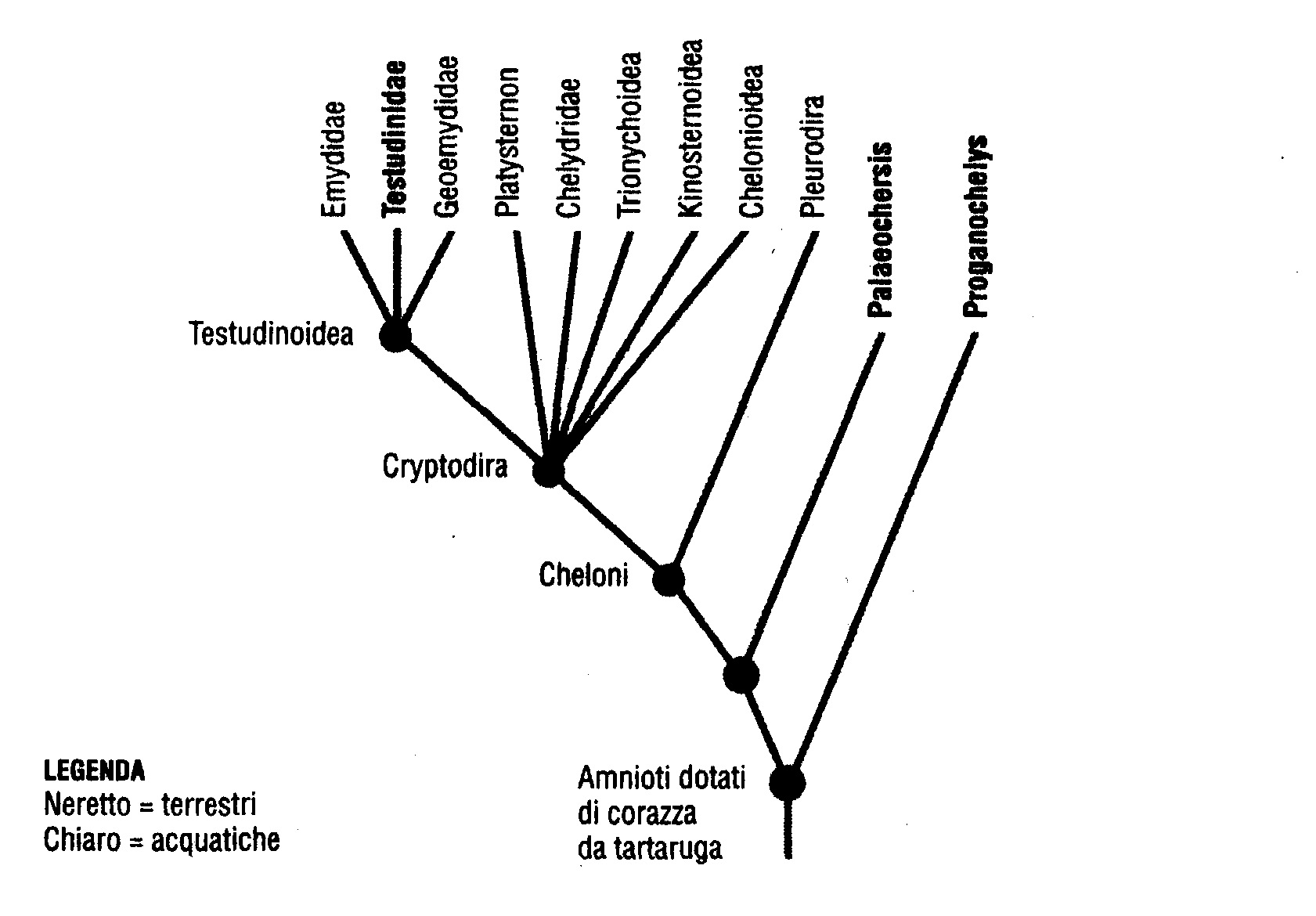
Nel diagramma le tartarughe terrestri sono rappresentate dalle scritte in neretto, e si vede che quelle odierne costituiscono un unico ramo, la famiglia Testudinidae, profondamente annidata in ricche ramificazioni di cheloni acquatici. Tutte le loro cugine strette sono marine. Le moderne tartarughe di terra sono un ramo di un cespuglio di tartarughe acquatiche, le cui antenate tornarono sulla terraferma. Il diagramma è compatibile con l'ipotesi che la corazza si sia evoluta in mare in una creatura come Odontochelys. Ma adesso abbiamo un'altra difficoltà. Se guardiamo l'albero filogenetico, noteremo che, oltre alle Testudinidae (tutte le tartarughe terrestri moderne), vi sono due generi fossili dotati di corazza completa: Proganochelys e Palaeochersis. Sono definiti terrestri per motivi che illustreremo tra poco, e si trovano fuori dei rami che rappresentano le tartarughe acquatiche.
Prima che fosse scoperto Odontochelys, i più antichi cheloni conosciuti erano Proganochelys e Palaeochersis. Come Odontochelys, vissero nel tardo Triassico, ma circa 15 milioni di anni dopo la tartaruga con mezza corazza. Alcuni biologi hanno ricostruito la loro storia definendoli cheloni di acqua dolce, ma prove recenti li pongono sulla terraferma, come indicato in neretto nel diagramma. Ci si potrà chiedere come facciamo a capire se animali fossili, di cui per giunta sono stati rinvenuti solo frammenti, vivessero sulla terraferma o in mare. A volte è del tutto evidente dove vivevano. Gli ittiosauri erano rettili contemporanei dei dinosauri, con pinne e corpo aerodinamico. I fossili sembrano delfini e, come i delfini, vivevano sicuramente in mare. Con le tartarughe la collocazione è meno ovvia. Comprensibilmente, l'indizio più chiaro è dato dagli arti. Le pinne natatorie sono assai diverse dalle zampe. Walter Joyce e Jacques Gauthier, della Yale University, sono partiti da questa semplice premessa per fornire le cifre che la confermano: Hanno preso tre misure chiave delle zampe anteriori e delle ossa della «mano» di settantuno specie di cheloni viventi. Resisterò alla tentazione di spiegare i loro eleganti calcoli, ma la conclusione che hanno tratto è stata chiara. Quegli animali avevano zampe per camminare, non pinne. Vivevano sulla terraferma. Erano però solo lontani cugini delle moderne tartarughe terrestri.
Sembra allora che abbiamo un problema. Se, come credono gli autori cinesi, Odontochelys dimostra che la mezza corazza si evolse in mare, come si spiegano i due generi terrestri Proganochelys e Palaeochersis, con corazza completa, di 15 milioni di anni dopo? Fino alla scoperta di Odontochelys, non avrei esitato a dire che Proganochelys e Palaeochersis erano rappresentanti del tipo ancestrale di terra prima del ritorno in mare. La corazza si evolse sulla terra. Alcune tartarughe con corazza tornarono in mare, come avrebbero fatto in seguito foche, balene e dugonghi, mentre altre rimasero sulla terraferma, ma si estinsero. Poi qualche tartaruga marina tornò sulla terraferma per dare origine a tutte le tartarughe terrestri moderne. Ecco che cosa avrei detto, e anzi che cosa di fatto avevo detto nella precedente stesura di questo capitolo, prima di apprendere che era stato scoperto Odontochelys. Ma Odontochelys ci riporta all'incertezza precedente. Adesso abbiamo tre possibilità, tutte equamente affascinanti.
1. Proganochelys e Palaeochersis sarebbero i sopravvissuti tra tutti gli animali terrestri che in precedenza avevano trasferito in mare alcuni loro rappresentanti, tra cui gli antenati di Odontochelys. Se così fosse, la corazza si sarebbe evoluta sulla terraferma e Odontochelys avrebbe perso il carapace in acqua, mantenendo solo il piastrone ventrale.
2. La corazza si sarebbe evoluta in mare, come suggerito dagli autori cinesi: il piastrone ventrale si sarebbe sviluppato per primo e il carapace dorsale in un secondo tempo. In tal caso che cosa dovremmo pensare di Proganochelys e Palaeochersis, vissuti sulla terraferma dopo che Odontochelys era vissuto in mare con la sua mezza corazza? Proganochelys e Palaeochersis potrebbero avere evoluto la corazza in maniera indipendente. Ma c'è una terza possibilità:
3. Proganochelys e Palaeochersis rappresenterebbero un precedente ritorno dal mare alla terraferma. Non è un'idea assolutamente entusiasmante?
Siamo già abbastanza sicuri che le tartarughe abbiano compiuto un'inversione di marcia dirigendosi verso la terraferma: un tipo iniziale di chelone terrestre tornò nell'ambiente marino dei suoi precedenti antenati pesci, divenne tartaruga marina, poi riapprodò sulla terra come nuova incarnazione di tartarughe terrestri, le Testudinidae. Questo lo sappiamo per certo o quasi per certo. Ma ora ci troviamo davanti all'ipotesi supplementare che il ritorno sia avvenuto due volte. Vi sarebbe stato non solo quello che diede origine alle tartarughe moderne, ma anche quello che molto prima, nel Triassico, aveva dato origine a Proganochelys e Palaeochersis.
In un altro libro ho definito il DNA il «Libro genetico dei morti» A causa del modo in cui funziona la selezione naturale, il DNA di un animale è in un certo senso la descrizione testuale dei mondi in cui gli antenati di quell'animale furono selezionati. Nel caso di un pesce, il libro genetico dei morti descrive i mari ancestrali. Nel caso dell'uomo e della maggior parte dei mammiferi, i primi capitoli sono tutti ambientati in mare e quelli successivi tutti sulla terraferma. Nel caso di balene, dugonghi, iguane marine, pinguini, foche, otarie e tartarughe, c'è una terza sezione del libro che racconta l'epico ritorno al banco di prova del remoto passato, il mare. Nel caso invece delle tartarughe terrestri, pare vi sia stato un doppio ritorno, avvenuto in maniera indipendente e in epoche ampiamente distanziate nel tempo; dunque c'è una quarta sezione del libro dedicata alla riemersione finale ‑ ma sarà davvero finale? ‑ verso la terraferma. Potrà mai esserci un altro animale per il quale il libro generico dei morti sia un tale palinsesto di multiple inversioni di marcia evolutive? Alla fine del capitolo non posso fare a meno di pormi qualche domanda sulle tartarughe d'acqua dolce e salmastra, che sono strette cugine delle tartarughe terrestri. Le loro antenate si trasferirono dal mare all'acqua salmastra e infine a quella dolce? Questo tipo di chelone costituisce forse uno stadio intermedio sulla via che dal mare va alla terraferma? O rappresenta un ennesimo ritorno all'acqua a partire da antenati che erano moderne tartarughe di terra? E se per tutto il tempo evolutivo i cheloni avessero fatto la spola tra mare e terraferma? Potrebbe, il palinsesto, essere stato riscritto ancor più volte di quelle che ho finora ipotizzato?
Il 19 maggio 2009, mentre correggevo le bozze di questo libro, la rivista scientifica on line «PLOS One» ha annunciato la scoperta dell'«anello mancante» tra primati simili a lemuri e primati simili a scimmie. Chiamato Darwinius masillae (ill. 9), l'esemplare visse 47 milioni di anni fa nella foresta pluviale di quella che oggi è la Germania. Gli autori dell'articolo affermano che è il fossile di primate più completo che sia mai stato rinvenuto: ci restano non solo le ossa, ma anche pelle, pelo, alcuni organi interni e il suo ultimo pasto. D. masillae è stato definito «l'ottava meraviglia del mondo» che «finalmente conferma la teoria evoluzionistica di Darwin», ma la vera meraviglia è la campagna pubblicitaria orchestrata in occasione della sua scoperta, con dovizia di slogan sensazionali come «la scoperta più importante degli ultimi 47 milioni di anni», un «avvenimento di risonanza mondiale» che «rivoluziona tutto», il «primo vero nesso con l'uomo», «immagini che una volta pubblicate avranno lo stesso impatto di un asteroide in rotta di collisione con la terra». Tutte assurdità; tuttavia è uno splendido fossile che getterà sicuramente luce sulle nostre ascendenze, e questo è un motivo sufficiente per mostrarlo in queste pagine. Mah, a quanto pare la stupida montatura dell'«anello mancante» non ha perso neanche un grammo del suo potere.
Nella sua opera più celebre, L'origine delle specie, Darwin dedica al tema dell'evoluzione umana solo undici portentose parole: «Verrà fatta luce sull'origine dell'uomo e sulla sua storia». Così si esprime nella prima edizione, quella che cito sempre, salvo diversa indicazione. Nella sesta (e ultima), Darwin si permise un rafforzativo e la frase diventò: «Molta luce verrà fatta sull'origine dell'uomo e sulla sua storia». Mi piace pensare al grande uomo che, con la penna sospesa sopra la quinta edizione, si chiede se sia il caso di concedersi il lusso dell'aggettivo «molta». Ma, anche così, la frase è fin troppo misurata.
Darwin decise di affrontare l'argomento dell'evoluzione umana in un altro libro, L'origine dell'uomo. Forse non stupisce che i due volumi originari di quell'opera più tarda dedichino più spazio al tema del sottotitolo, «e la scelta sessuale» (analizzata soprattutto negli uccelli), che all'evoluzione umana. Non stupisce perché, all'epoca in cui Darwin scriveva, non c'era nessun fossile che ci collegasse con i primati, nostri parenti più stretti. Darwin disponeva soltanto di primati viventi da osservare e se ne servì nel modo giusto: sostenne correttamente (e pressoché da solo) che i nostri più stretti cugini viventi erano tutti africani (gorilla e scimpanzé: all'epoca non si distinguevano i bonobi dagli scimpanzé, ma in ogni caso anche i bonobi sono africani) e predisse quindi che, per trovare fossili umani ancestrali, si sarebbe dovuto cercare in Africa. Si rammaricava della scarsità di fossili, ma era ottimista. Citando Lyell, il celebre geologo dell'epoca divenuto suo mentore, osservava che «in tutte le classi di vertebrati la scoperta di resti fossili è stata un processo particolarmente lento e fortuito» e aggiungeva: «Non si dimentichi che le regioni dove è più probabile reperire i resti che collegano l'uomo con una creatura scimmiesca estinta non sono ancora state perlustrate dai geologi». Si riferiva all'Africa, e non fu d'aiuto che i suoi immediati successori abbiano quasi completamente ignorato i suoi consigli e cercato invece in Asia.
Ciò nonostante, in Asia gli «anelli mancanti» cominciarono a mancare sempre meno. I primi fossili scoperti erano relativamente recenti: avendo meno di un milione di anni, risalivano a un'epoca in cui gli ominidi erano abbastanza simili agli esseri umani moderni ed erano emigrati dall'Africa per raggiungere l'Estremo Oriente. Furono chiamati «Uomo di Giava» e «Uomo di Pechino» dai luoghi in cui vennero rinvenuti. L'Uomo di Giava fu scoperto dall'antropologo olandese Eugene Dubois nel 1891. Dubois lo chiamò Pithecanthropus erectus, cioè uomo‑scimmia, ritenendo di aver trovato l'«anello mancante». Si levarono due critiche contrastanti, che perciò suonavano piuttosto come conferme: alcuni dissero che il fossile era esclusivamente umano, altri che era un gibbone gigante. In seguito, nella sua vita funestata da rancori e litigiosità, quando furono scoperti i fossili di Pechino e qualcuno osò dire che somigliavano al suo reperto, per proteggere la sua creatura e riconfermarne il valore come l'unico vero anello mancante, definì i reperti di Uomo di Pechino molto più vicini all'uomo moderno e sostenne che l'Uomo di Giava del giacimento di Trinil era intermedio tra le scimmie antropomorfe e l'uomo.
Pithecanthropus non era un uomo, ma un genere gigantesco simile ai gibboni, anche se era superiore ai gibboni per il cervello molto grande e per la sua capacità di assumere stazione e andatura erette. Aveva una cefalizzazione [rapporto tra dimensioni del cervello e dimensioni del corpo} che era il doppio di quella delle scimmie antropoidi e la metà di quella dell'uomo...
E’ stato il sorprendente volume del cervello, troppo grande rispetto a quello di una scimmia antropoide e tuttavia piccolo in confronto alla media umana (benché non più piccolo del più piccolo cervello umano), a produrre oggi il generale convincimento che l'«Uomoscimmia» di Trinil, l'Uomo di Giava, fosse effettivamente un uomo primitivo. Dal punto di vista morfologico, però, la calotta cranica assomiglia molto a quella delle scimmie antropoidi, in particolare a quella del gibbone ...
Non deve avere migliorato il temperamento irascibile di Dubois vedere che il suo discorso era stato frainteso e che gli altri avevano creduto dicesse che Pithecanthropus era solo un gibbone gigantesco, non uno stadio intermedio tra il gibbone e l'uomo. Non à caso si premurò di ribadire quanto aveva affermato in precedenza: «Continuo a credere, più fermamente che mai, che il Pithecanthropus di Trinil sia il vero "anello mancante"».
Ogni tanto i creazionisti hanno usato come arma politica la notizia secondo la quale Dubois avrebbe fatto a un certo punto marcia indietro rispetto alla dichiarazione iniziale con la quale aveva definito Pithecanthropus un uomo‑scimmia intermedio. L'associazione creazionista Answers in Genesis ha però aggiunto la storia di Dubois alla sua lista degli argomenti screditati da non utilizzare. Torna a suo merito che l'argomento permanga nella lista. Come ho detto, da studi recenti sia l'esemplare di Giava sia l'esemplare di Pechino di Pithecanthropus sono risultati abbastanza giovani: meno di un milione di anni. Ora si ritiene appartengano, come noi, al genere Homo, e hanno mantenuto il nome specifico di erectus che aveva dato loro Dubois: Homo erectus.
Dubois scelse la parte sbagliata di mondo per la sua ossessiva ricerca dell'«anello mancante». Era naturale che un olandese si dirigesse per prima cosa nelle Indie Orientali olandesi, ma un uomo dotato come lui di tanta passione naturalistica avrebbe dovuto seguire il consiglio di Darwin e andare in Africa; perché, come vedremo, è in Africa che si sono evoluti i nostri antenati. Allora che cosa ci facevano quegli esemplari di Homo erectus fuori dell'Africa?* È importante ricordare che vi furono due esodi, che non bisogna confondere. Abbastanza di recente, forse meno di 100.000 anni fa, gruppi sparsi di Homo sapiens dall'aspetto molto simile al nostro lasciarono l'Africa e si diversificarono nelle varie razze che vediamo oggi nel mondo: inuit, nativi americani, nativi australiani, cinesi e così via. Ma ci fu un esodo precedente dall'Africa, durante il quale i pionieri erectus lasciarono in Asia ed Europa dei fossili, compresi gli esemplari di Giava e Pechino. Il fossile non africano più antico è stato rinvenuto in un paese dell'Asia centrale, la Georgia, ed è stato chiamato Homo georgicus (fig. 7.1).
Era una piccola creatura il cui cranio (abbastanza ben conservato), secondo gli odierni metodi di calcolo, risale a circa 1,8 milioni di anni fa. Alcuni tassonomisti (anche se altri non lo riconoscono come specie a sé) lo hanno chiamato Homo georgicus per indicare che è più primitivo del resto dei primi profughi dell'Africa, i quali sono tutti classificati come Homo erectus. Arnesi di pietra leggermente più vecchi di Homo georgicus sono appena stati scoperti in Malaysia e hanno scatenato una nuova ricerca di ossa fossili in quella penisola. In ogni caso, i più antichi fossili asiatici sono abbastanza vicini all'uomo moderno e oggi sono tutti classificati come Homo; per trovare loro predecessori più anziani bisogna andare in Africa. Prima, però, facciamo una pausa per chiederci che cosa è lecito aspettarsi da un «anello mancante'>.
Supponiamo per ipotesi di prendere sul serio l'espressione «anello mancante» e di cercare un intermedio tra gli scimpanzé e noi. Noi non discendiamo dagli scimpanzé, ma è molto probabile che il nostro antenato comune somigliasse più a loro che a noi: non aveva un cervello voluminoso come il nostro, quasi sicuramente non camminava eretto, era molto più peloso di noi e non aveva certo caratteristiche umane complesse come il linguaggio. Così, sebbene dobbiamo sostenere fermamente, davanti al comune malinteso, che non discendiamo dagli scimpanzé, non c'è niente di male se ci chiediamo quale aspetto avrebbe uno stadio intermedio tra uno scimpanzé e noi.
Pelo e linguaggio non si fossilizzano bene, ma possiamo procurarci buoni indizi sulle dimensioni del cervello osservando il cranio, e buoni indizi sull'andatura esaminando l'intero scheletro (compreso il cranio, perché il foramen magnum, il foro occipitale dal quale passa la colonna vertebrale, è orientato verso il basso nei bipedi, più in senso posteriore nei quadrupedi). I possibili candidati alla qualifica di anello mancante potrebbero avere uno dei seguenti attributi:
1. Dimensioni del cervello e andatura intermedi: magari un incedere curvo e dinoccolato anziché il fiero portamento eretto tanto amato dai sergenti maggiori e dalle insegnanti delle scuole femminili di una volta.
2. Cervello delle dimensioni di quello dello scimpanzé, con stazione eretta umana.
3. Cervello grande, più simile a quello umano, con andatura quadrupede come quella dello scimpanzé.
Tenendo presenti queste tre possibilità, esaminiamo alcuni dei molti fossili africani di cui disponiamo adesso, ma che purtroppo non erano stati ancora scoperti all'epoca di Darwin.
Le prove molecolari (su cui tornerò nel X capitolo) dimostrano che l'antenato da noi condiviso con gli scimpanzé visse circa sei milioni di anni fa o ancora prima; dividiamo quindi a metà questo arco di tempo e cerchiamo fossili di circa tre milioni di anni. Il fossile più famoso di questo periodo è «Lucy», chiamata da Donald Johanson, che la scoprì in Etiopia, Australopithecus afarensis. Purtroppo abbiamo solo frammenti del suo cranio, ma la mandibola è insolitamente ben conservata. Lucy era piccola rispetto ai parametri moderni, anche se non piccola come Homofloresiensis, la minuscola creatura che la stampa ha avuto il cattivo gusto di soprannominare «lo Hobbit», e che si estinse in tempi incredibilmente recenti sull'Isola di Flores, in Indonesia. Lo scheletro di Lucy è abbastanza completo da permetterci di capire che camminava eretta sul terreno, ma probabilmente cercava anche rifugio tra gli alberi, dove era un'agile arrampicatrice. Prove convincenti dimostrano che le ossa attribuitele sono appartenute davvero a un unico individuo. Lo stesso non vale per la cosiddetta «Prima famiglia», rinvenuta sempre in Etiopia e composta da ossa di almeno tredici individui simili a Lucy, i quali, circa alla sua stessa epoca, rimasero in qualche modo seppelliti insieme. I frammenti di Lucy e della Prima famiglia consentono di farsi un'idea dell'aspetto di Australopithecus afarensis, ma è difficile procedere a una ricostruzione completa a partire da reperti di molti individui. Per fortuna, un cranio abbastanza completo, chiamato AL 444‑2 (fig. 7.3), fu scoperto nel 1992 nella stessa regione dell'Etiopia, l'Afar, e il suo esame ha confermato la ricostruzione che era stata fatta in precedenza.
Dall'analisi di Lucy e dei suoi simili si è concluso che gli A. afarensis avevano il cervello delle stesse dimensioni di quello degli scimpanzé, ma, diversamente dagli scimpanzé, camminavano eretti sulle zampe posteriori come noi, il che corrisponde allo scenario ipotetico n. 2. «Le Lucy» erano un po' come scimpanzé bipedi. La loro stazione eretta è stata clamorosamente confermata dalla straordinaria serie di orme fossilizzate nella cenere vulcanica, scoperta da Mary Leakey. Le orme si trovano più a sud, a Laetoli, in Tanzania, e sono più antiche di Lucy e di AL 444‑2: risalgono a 3,6 milioni di anni fa. Sono in genere attribuite a due Australopithecus afarensis che camminavano insieme (mano nella mano?), ma l'importante è che, 3,6 milioni di anni fa, una scimmia antropomorfa camminava eretta sulla terra, con piedi che somigliavano molto ai nostri, anche se il cervello era come quello di uno scimpanzé.
È molto probabile che la specie da noi denominata Australopithecus afarensis, la specie di Lucy, comprendesse i nostri antenati di tre milioni di anni fa. Altri fossili sono stati attribuiti a specie diverse dello stesso genere ed è praticamente certo che i nostri antenati fossero membri di quel genere. Il primo esemplare di Australopitecine mai scoperto, l'olotipo del genere Australopithecus, è il cosiddetto bambino di Taung. All'età di tre anni e mezzo, il bambino di Taung fu divorato da un'aquila. Lo dimostrano i segni delle ferite alle orbite del fossile, identici a quelli che le aquile moderne lasciano alle scimmie moderne quando strappano loro gli occhi. Povero piccino di Taung, che urlavi al vento mentre l'aquila, nella sua furia, ti sollevava in alto! Non ti avrebbe certo consolato sapere che due milioni e mezzo di anni dopo saresti diventato famoso come olotipo di Australopithecus africanus. Povera mamma di Taung, che piangesti nel Pliocene.
L'olotipo è il primo individuo di una nuova specie a essere descritto e a ricevere ufficialmente la targhetta vergine nel museo. Di norma, è il termine di confronto dei successivi reperti per vedere se corrispondono. A scoprire il bambino di Taung e a dargli una nuova nomenclatura binomia (genere e specie) fu l'antropologo sudafricano Raymond Dart, nel 1924.
Che differenza c'è tra «specie» e «genere»? Prima di proseguire diamo subito una risposta a questa domanda. Il genere è la categoria più inclusiva e spesso comprende più specie; la specie rientra nel genere. Homo sapiens e Homo erectus sono due specie all'interno del genere Homo. Australopithecus africanus e Australopithecus afarensis sono due specie all'interno dello stesso genere, Australopithecus. Il nome scientifico latino di un animale o una pianta è sempre costituito dal nome generico, cioè del genere (maiuscolo), seguito dal nome specifico, cioè della specie (minuscolo). Entrambi i nomi sono scritti in corsivo. A volte esiste anche il nome subspecifico, della sottospecie, il quale segue quello specifico, come in Homo sapiens neandertalensis. I tassonomisti spesso litigano sui nomi. Molti, per esempio, vorrebbero si dicesse Homo neandertalensis anziché Homo sapiens neandertalensis, una nomenclatura che eleva l'uomo di Neandertal dallo stato di sottospecie a quello di specie. Spesso non si è d'accordo nemmeno sui nomi generici e i nomi specifici, che a volte vengono cambiati in seguito a revisioni della letteratura scientifica. Paranthropus boisei alla sua epoca era Zinjanthropus boisei e Australopithecus boisei,* e ancora oggi lo si definisce sovente, in maniera informale, Australophitecus robustus, che si contrappone alle succitate specie di gracile. Uno dei principali messaggi di questo capitolo riguarda la natura piuttosto arbitraria della classificazione zoologica.
Raymond Dart, dunque, diede il nome di Australopithecus al bambino di Taung, l'olotipo del genere, e da allora ci è toccato sopportare che il nostro progenitore portasse questo nome terribilmente prosaico. Un nome che significa soltanto «scimmia meridionale». Dart avrebbe ben potuto inventare un nome più fantasioso per un genere così importante; avrebbe potuto perfino intuire che altri membri di quel genere sarebbero stati in seguito scoperti a nord dell'equatore.
Leggermente più vecchia del bambino di Taung è Mrs Pies (fig. 7.4.), uno dei crani meglio conservati che abbiamo, anche se privo di mandibola.
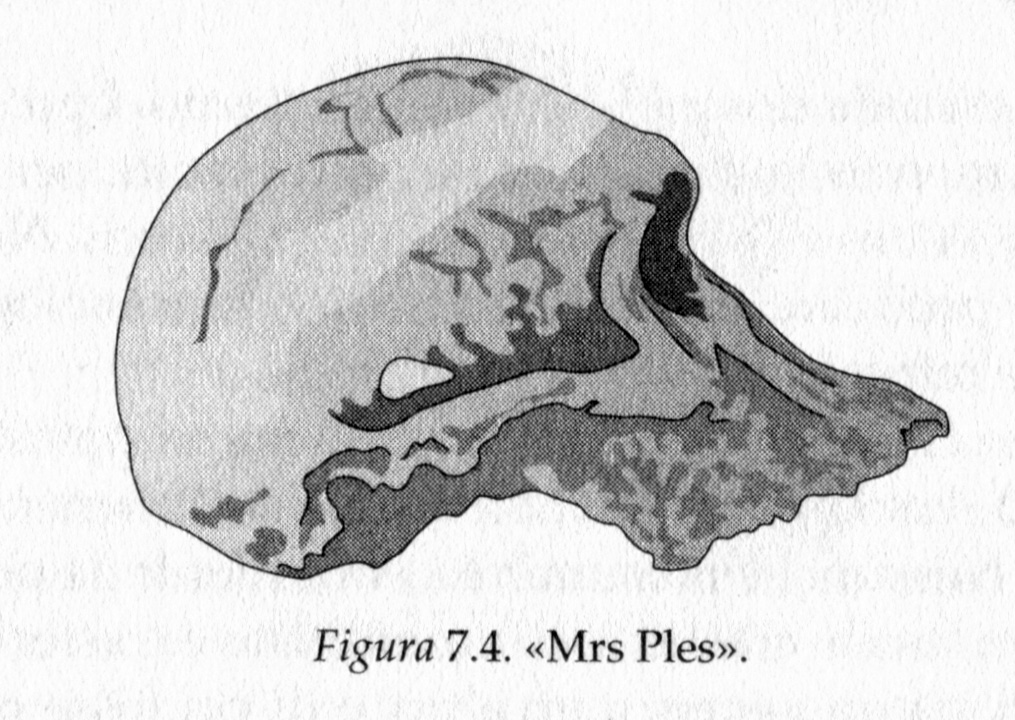
Mrs Ples, che in realtà doveva essere un piccolo maschio anziché una grossa femmina, fu chiamata così perché in origine venne assegnata ai genere Plesianthropus, nome che significa «quasi umano» e che è sicuramente meglio di «scimmia meridionale». Era lecito sperare che, quando successivi tassonomisti decisero di assegnare Mrs Ples e i suoi simili allo stesso genere del bambino di Taung, Plesianthropus diventasse il nome di tutti loro, ma purtroppo le regole della nomenclatura zoologica sono così rigide da sfiorare la pedanteria. Una volta dato un nome, il diritto di precedenza ha la meglio sul buonsenso e la logica. «Scimmia meridionale» sarà anche una denominazione orrenda, ma non importa: è stato dato prima del molto più ragionevole Plesianthropus e ci tocca sorbircelo, a meno che... Io continuo a sperare malignamente che qualcuno scopra, nel cassetto impolverato di un museo sudafricano, un fossile da tempo dimenticato, appartenente senza ombra di dubbio ai genere di Mrs Ples e del bambino di Taung, ma recante un'etichetta con la scritta «olotipo di Hemianthropus, 1920». Di colpo tutti i musei del mondo sarebbero costretti a rinominare i loro esemplari e calchi di Australopithecus, e tutti i libri e gli articoli sulla preistoria ominide dovrebbero fare altrettanto. I programmi informatici del mondo lavorerebbero giorno e notte per eliminare i vari Australopithecus e sostituirli con Hemianthropus. Non vedo altro modo per produrre un cambiamento di terminologia repentino, generale e retroattivo.
Ora è chiaro che quando Mrs Ples mutò nome, passando da Plesianthropus ad Australopithecus, non mutò assolutamente nulla nel mondo reale. Nessuno, è presumibile, ebbe niente da obiettare. Ma pensiamo a un fossile che venisse riesaminato e trasferito, per motivi anatomici, da un genere a un altro, o di cui fosse messo in discussione ii genere (cosa che accade assai di frequente) da antropologi rivali. Dopotutto, per la logica dell'evoluzione è essenziale che siano esistiti individui posti esattamente sulla linea di confine tra due generi, poniamo Australopithecus e Homo. È facile guardare Mrs Pies e l'Homo sapiens moderno e dire, sì, non vi è dubbio che i due crani appartengano a generi diversi. Se assumiamo, come fa la stragrande maggioranza degli antropologi odierni, che tutti i membri del genere Homo discendano da antenati appartenenti al genere denominato Australopithecus, a un certo punto, lungo la catena della discendenza di una specie dall'altra, è inevitabile vi sia stato almeno un individuo che si collocava esattamente sulla linea di confine. E’ un concetto importante, quindi mi ci soffermerò ancora un poco.
Tenendo presente la forma del cranio di Mrs Pies e ricordando che rappresenta un Australopithecus africanus di 2,6 milioni di anni fa, si osservino i crani di KNM ER 1813 e di KNM ER 1470 (figg. 7.5 e 7.6).
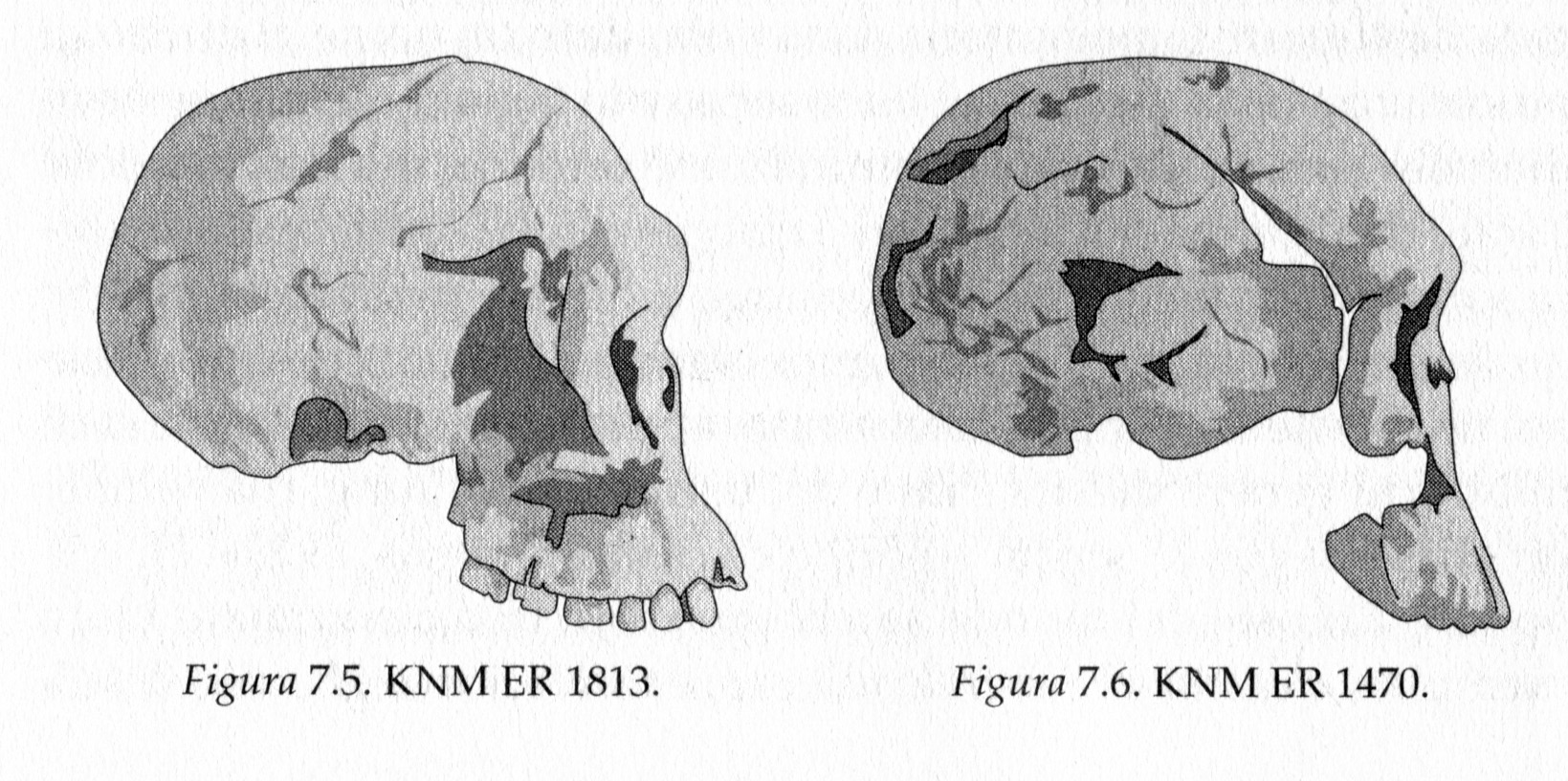
I due reperti risalgono a circa 1,9 milioni di anni fa e sono stati posti nel genere Homo dalla maggior parte degli esperti. Oggi 1813 è classificato come Homo habilis, ma non è stato sempre così. Fino a poco tempo fa anche 1470 era ritenuto un habilis, ma oggi c'è chi si sta adoperando per riclassificarlo come Homo rudolfensis. Ancora una volta vediamo quanto instabili e transitori siano i nomi. Ma non importa: i due fossili hanno, per comune accordo degli esperti, un piede nel genere Homo. La differenza evidente rispetto a Mrs Ples e ai suoi simili è che questi avevano il viso più prognato e una scatola cranica più piccola. Sotto entrambi questi profili, 1813 e 1470 appaiono più umani e Mrs Ples risulta più «scimmiesca».
Ora guardiamo il cranio chiamato Twiggy (fig. 7.7).
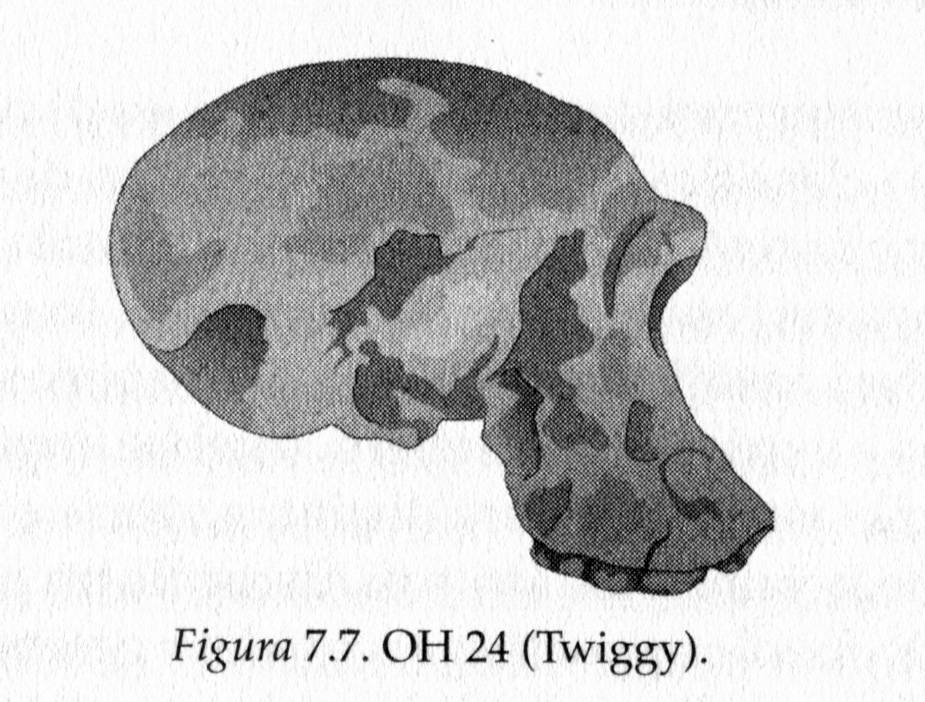
Anche Twiggy oggi è generalmente classificata come Homo habilis. Ma il viso prognato ricorda più quello di Mrs Ples che quello di 1470 e 1813. Forse non vi sorprenderà sapere che Twiggy è stata posta da alcuni antropologi nel genere Australopithecus e da altri nel genere Homo. Di fatto, ognuno di questi tre fossili è stato classificato, a seconda dei periodi, Homo habilis o Australopithecus habilis. Come ho già spiegato, a seconda dei momenti alcuni paleoantropologi hanno dato a 1470 un nome specifico diverso, convertendo habilis in Rudolfensis. Per giunta, il nome specifico rudolfensis è stato associato sia al nome generico Australopithecus sia al nome generico Homo. In sintesi, i tre fossili sono stati denominati in vario modo da esperti diversi in periodi diversi,6 e precisamente:
KNM ER 1813 Australopithecus habilis, Homo habilis
KNM ER 1470 Australopithecus habilis, Homo habilis, Australopithecus rudolfensis, Homo rudolfensis
0H24 (Twiggy) Australopithecus habilis, Homo habilis
Una simile confusione terminologica non dovrebbe incrinare la nostra fiducia nella biologia evoluzionistica? Nient'affatto; semmai il contrario. La confusione è inevitabile, in quanto tutte quelle creature sono stadi intermedi evolutivi, anelli che un tempo mancavano ma ora non mancano più. Dovremmo preoccuparci molto se non ci fossero intermedi così vicini alle linee di confine da risultare di difficile classificazione. Anzi, dal punto di vista evolutivo conferire nomi distinti diventerebbe praticamente impossibile se la documentazione fossile fosse più completa. In un certo senso è una fortuna che i fossili siano così rari. Se disponessimo di una riserva continua e ininterrotta di reperti, sarebbe impossibile o almeno assai difficile assegnare nomi distinti a specie e generi. E' lecito concludere che la principale fonte di discordia tra paleoantropologi ‑ se un certo fossile appartenga a un dato genere/specie piuttosto che a un altro ‑ di una futilità assoluta quanto significativa.
Supponiamo che per un caso fortunato, abbiamo una documentazione fossile continua di tutti i cambiamenti evolutivi, senza alcun anello mancante. Ora guardiamo i quattro nomi latini che sono stati attribuiti a 1470. A prima vista il passaggio da habilis a rudolfensis parrebbe più insignificante del passaggio da Australopithecus a Homo. Due specie all'interno di un genere si assomigliano più di due generi, no? Non è questo il succo della distinzione tra un genere e l'altro (come Homo e Pan, due generi diversi di ominidi africani) e tra una specie e l'altra (come troglodytes e paniscus, due specie diverse del genere scimpanzé) nella gerarchia sistematica? Sì, questo è vero quando classifichiamo gli animali moderni, che si possono considerare le punte dei rami dell'albero evolutivo, con gli antecedenti all'interno della corona dell'albero adeguatamente morti e scomparsi. Naturalmente, i rami che si uniscono più indietro (sempre all'interno della corona dell'albero) saranno meno simili di quelli la cui connessione (l'antenato comune più recente) è più vicina alle punte. Il sistema funziona purché non cerchiamo di classificare gli antecedenti morti. Ma ove includessimo la documentazione fossile ipoteticamente completa, le distinzioni nette verrebbero a mancare e, come regola generale, non si potrebbero più usare nomi distinti. Lo vediamo con chiarezza se proviamo ad andare costantemente a ritroso nel tempo, come abbiamo fatto con i conigli del II capitolo.
A mano a mano che risaliamo alle origini dell'Homo sapiens moderno, arriverà il momento in cui la differenza rispetto alle persone viventi diventerà così grande da indurci a dare un nome specifico diverso da sapiens, e cioè Homo ergaster. Tuttavia, lungo questa strada a ritroso, gli individui devono essere stati a ogni passo abbastanza simili ai propri genitori e ai propri figli da essere collocati nella medesima specie. Ora, se risaliamo ancora più indietro nel tempo alla ricerca delle origini di Homo ergaster, verrà il momento in cui raggiungeremo individui abbastanza diversi dall'ergaster «comunemente accettato» da meritare un nome specifico diverso, diciamo Homo habilis.
Veniamo ora al nocciolo della questione. Procedendo sempre a ritroso, a un certo punto incontreremo inevitabilmente individui abbastanza diversi dall'Homo sapiens moderno da meritare un nome generico diverso, diciamo Australopithecus. Il guaio è che dire «abbastanza diversi dall'Homo sapiens moderno» è tutt'altro che dire «abbastanza diversi dal primo Homo», qui definito Homo habilis. Riflettiamo sulla nascita del primo esemplare di Homo habilis. I suoi genitori erano degli Australopithecus. Allora apparteneva a un genere diverso da quello di suo padre e sua madre? Ma è un'assurdità! Sì, infatti; tuttavia non è la realtà a essere in difetto, bensì la pervicace volontà umana di assegnare ogni cosa a una categoria dotata di un nome. Nella realtà non c'è mai stata una creatura come il primo esemplare di Homo habilis. Non è esistito un primo esemplare di nessuna specie, nessun genere, nessun ordine, nessuna classe e nessun phylum. Ogni creatura venuta al mondo, se ci fosse stato nei pressi uno zoologo pronto a classificarla, sarebbe stata classificata come appartenente alla stessa specie dei suoi genitori e dei suoi figli. Tuttavia, con il senno di poi della modernità e con il vantaggio ‑ sì, in questo senso paradossale possiamo chiamarlo un vantaggio ‑ della mancanza della maggior parte degli anelli, diventa possibile la suddivisione in specie, generi, famiglie, ordini, classi e phyla distinti.
Vorrei che disponessimo davvero di una lunga serie completa e ininterrotta di fossili, una documentazione cinematica di tutti i cambiamenti evolutivi avvenuti. Lo vorrei, anche perché mi piacerebbe veder rimanere con un palmo di naso tutti quegli zoologi e antropologimche passano la vita a litigare tra loro su questioni di lana caprina, come se un certo fossile appartenga a una specie piuttosto che a un'altra, a un genere piuttosto che a un altro. Signori (mi chiedo perché non siano mai signore), state discutendo di parole, non di realtà. Come disse lo stesso Darwin nell'Origine dell'uomo: «In una serie di forme che derivano gradualmente da qualche creatura simile alle scimmie fino all'uomo come ora esiste, sarebbe impossibile definire il punto in cui si dovrebbe usare il vocabolo uomo».
Proseguiamo la nostra analisi dei fossili guardando anelli più recenti, tra quelli che mancavano all’epoca di Darwin e ora non mancano più. Quali intermedi troviamo tra noi e creature come 1470 e Twiggy, chiamate a volte Homo e a volte Australopithecus? Ne abbiamo già incontrati alcuni, come l'Uomo di Giava e l'Uomo di Pechino, in genere classificati come Homo erectus (fig. 7.8), ma entrambi vivevano in Asia, mentre da numerose prove risulta che la maggior parte dell'evoluzione umana si è svolta in Africa.
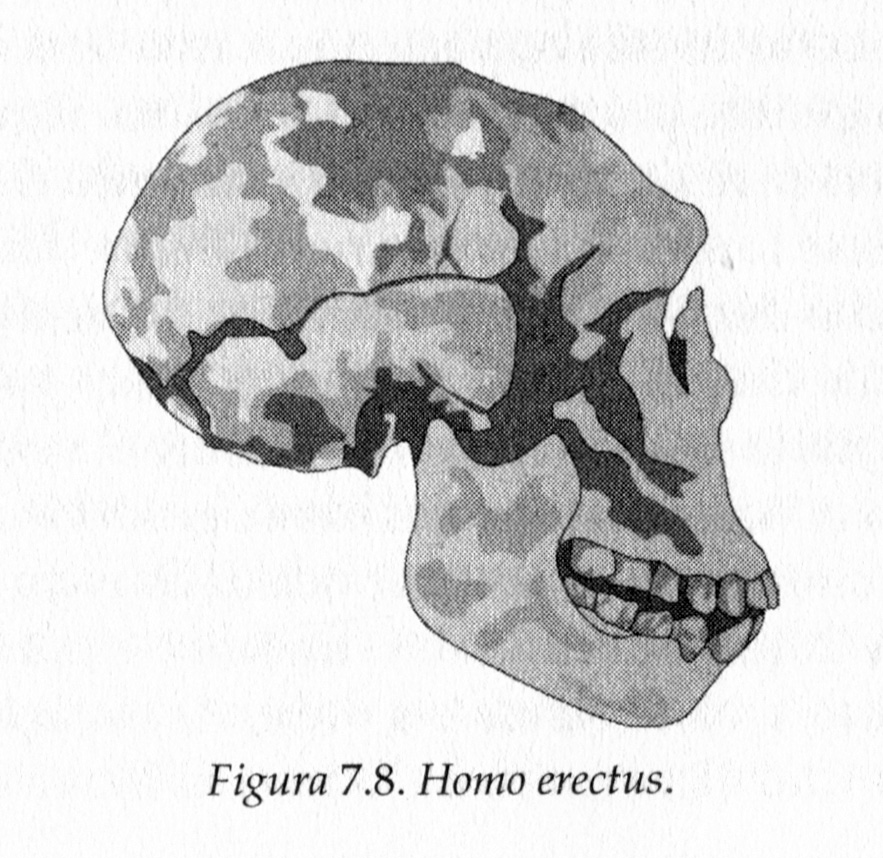
L'Uomo di Giava, l'Uomo di Pechino e i loro simili migrarono dalla loro patria africana. All'interno dell'Africa, i loro omologhi sono oggi classificati di solito come Homo ergaster, anche se per molti anni sono stati chiamati Homo erectus, a riprova dell'instabilità delle nostre procedure di classificazione. L'esempio più famoso di Homo ergaster, e uno dei fossili preumani più completi che siano mai stati trovati, è il ragazzo del Turkana o ragazzo d ariokotome, rinvenuto da Kamoya Kimeu, grande scopritore di ossili dell'équipe di paleontologi guidata da Richard Leakey.
Il ragazzo del Turkana visse circa 1,6 milioni di anni fa e morì a circa undici anni. Da alcuni indizi è lecito dedurre che se fosse arrivato all'età adulta avrebbe raggiunto la statura approssimativa di un metro e ottanta. Secondo le proiezioni, il volume del suo cervello adulto sarebbe stato di 900 centimetri cubi (cc). Era un tipico cervello da Homo ergaster/erectus, che si aggirava sui 1000 cc: notevolmente più piccolo di quello dell'uomo moderno, che oscilla tra i 1300 e i 1400 cc, ma più grande di quello di Homo habilis (circa 600 cc), a sua volta maggiore di quelli di Australopithecus (400 cc) e degli scimpanzé (circa lo stesso). Come si ricorderà, abbiamo concluso che il nostro antenato di tre milioni di anni fa aveva il cervello di uno scimpanzé, ma camminava eretto. Da questo potremmo arguire che la seconda metà della storia evolutiva, da tre milioni di anni fa a tempì recenti, sia stata contrassegnata dall'incremento progressivo delle dimensioni del cervello; e in effetti così pare sia stato.
Homo ergaster/erectus, di cui abbiamo molti esemplari fossili, è un anello assai convincente e non più mancante, a metà strada tra l'Homo sapiens moderno e l'Homo habilis di tre milioni di anni fa, che come abbiamo visto poteva essere definito a tutti gli effetti uno scimpanzé dalla stazione eretta. Quanti anelli bisogna avere prima che si ammetta che non sono più «mancanti»? E si può colmare anche il divario tra l'Homo ergaster e l'Homo sapiens moderno? Sì, abbiamo a disposizione una ricca riserva di fossili intermedi che coprono le ultime centinaia di migliaia di anni. Ad alcuni sono stati dati nomi di specie, come Homo heidelbergensis, Homo rhodesiensjs e Homo neandertalensis; altri (e a volte gli stessi) sono definiti Homo sapiens «arcaici». Ma, ribadisco, i nomi non contano. Quello che conta è che gli anelli non sono più mancanti. Gli stadi intermedi abbondano.
Vada a vedere al museo
Disponiamo dunque di una bella documentazione fossile sul cambiamento graduale che da Lucy, lo «scimpanzé dalla stazione eretta» di tre milioni di anni fa, porta a noi, all'umanità moderna. Come reagiscono i negazionisti davanti a questa evidenza? Alcuni, appunto, negando. Me ne sono reso conto nel corso di un'intervista che ho condotto nel 2008 per il documentario di Channel Four The Genius of Charles Darwin. L'intervistata era Wendy Wright, presidente del Concerned Women for America. Avendo dichiarato che «la pillola del giorno dopo è la migliore amica del pedofilo», la Wright aveva già dato discreta prova del suo raziocinio e nel corso dell'intervista ha confermato in pieno le attese. Nel documentario è andata in onda solo una minima parte dell'intervista. Quella che segue è una trascrizione molto più ampia del nostro dialogo, ma siccome naturalmente la riporto in relazione all'argomento che sto trattando, mi limito a citare i punti in cui abbiamo discusso della documentazione storica sulle ascendenze umane.
Wendy: Mi preme sottolineare che gli evoluzionisti non hanno il sostegno di dati scientifici. Eppure succede che la scienza che non sostiene l'evoluzione venga censurata. Per esempio non si dice che non esistono prove di un passaggio da una specie all'altra. Se ci fosse stata evoluzione, se si fosse passati dagli uccelli ai mammiferi o... o ancora oltre, ci sarebbe almeno qualche prova.
Richard: Ci sono innumerevoli prove. Purtroppo, voi continuate a ripetere questa sorta di mantra perché parlate solo tra di voi e ascoltate solo voi stessi. Se vi decideste una buona volta ad aprire gli occhi e a esaminare le prove...
Wendy: Me le mostri, me le mostri. Mi mostri le ossa, mi mostri le carcasse, mi mostri le prove degli stadi intermedi tra una specie e l'altra.
Richard: Ogni volta che si scopre un fossile intermedio tra una specie e l'altra, voi dite: «Ah, adesso abbiamo due lacune dove prima ce n'era una sola». Sa, quasi ogni fossile che viene scoperto è intermedio tra qualcosa e qualcos'altro.
Wendy [ride J: Se così fosse, lo Smithan Natural History Museum sarebbe pieno di questi esemplari, ma non lo è.
Richard: Lo è, lo è eccome. Nel caso degli esseri umani, dall'epoca di Darwin si è accumulata una quantità enorme di prove fossili riguardanti gli stadi intermedi. Abbiamo per esempio varie specie di Australopithecus e...poi c'è Homo habilis, che è intermedio tra Australopithecus, una specie più antica, e Homo sapiens, una specie più recente. Si può sapere perché non li considerate degli intermedi?
Wendy: ... Se a favore dell'evoluzione ci fossero state prove autentiche, sarebbero esposte nei musei, non solo nelle illustrazioni.
Richard: Le ho appena parlato di Australopithecus, Homo habilis, Homo erectus, Homo sapiens, Homo sapiens arcaico e Homo sapiens moderno: è una bella serie di intermedi.
Wendy: Non avete ancora le prove materiali, per cui...
Richard: Le prove materiali ci sono eccome. Vada a vedere al museo... Naturalmente non le ho qui con me, ma può entrare in qualsiasi museo e vedere Australopithecus, Homo habilis, Homo erectus, Homo sapiens arcaico e Homo sapiens moderno. Una bella serie di intermedi. Perché continua a dire «Mi mostri le prove>»quando gliele ho ripetutamente indicate? Vada a vedere al museo.
Wendy: Ci sono andata. Sono andata nei musei, ma siamo in molti a non essere ancora convinti.
Richard: Ha visto Homo erectus?
Wendy: A me pare che cerchiate in tutti i modi, aggressivamente, di farci cambiare idea e censurarci. Fate così perché siete frustrati dal fatto che tante persone continuino a non credere all'evoluzione. Se gli evoluzionisti fossero così sicuri delle loro idee, non cercherebbero in tutti i modi di censurare le nostre informazioni. Questo dimostra che l'evoluzione è ancora carente e controversa.
Richard: Sono.., ammetto di essere frustrato. Non per lo scetticismo della gente, ma perché le ho nominato quattro o cinque fossili... [Wendy ride e lei fa finta di non aver sentito.] Perché non va a guardarli?
Wendy: ... Se fossero nei musei che ho visitato tante volte, li guarderei con spirito imparziale, ma mi preme sottolineare...
Richard: I fossili sono al museo.
Wendy: Mi preme sottolineare che la filosofia evoluzionistica conduce a ideologie che si sono rivelate molto distruttive per la razza umana.
Richard: Sì, ma non sarebbe una buona idea se, invece di puntare il dito sulle sue cattive interpretazioni, che sono state usate a tarpi fini politici, lei cercasse di capire il darwinismo? Se lo facesse, sarebbe in grado di dimostrare l'infondatezza di quegli orribili travisamenti.
Wendy: Mah, in realtà subiamo di continuo un vero e proprio assalto da parte dei fautori dell'evoluzione. Non è che ci manchino le informazioni che continuate a presentarci. Non è che non le conosciamo, perché ci vengono propinate ossessivamente e non possiamo fare a meno di sentirle. Ma credo siate frustrati perché tanti di noi, pur avendo visto i vostri dati, continuano a non aderire alla vostra ideologia.
Richard: Ha visto Homo erectus? Ha visto Homo habilis? Ha visto Australopithecus? Questo le avevo chiesto.
Wendy: Ho visto una cosa: nei musei e nei manuali, ovunque affermino di mostrare le differenze evolutive tra una specie e l'altra, presentano solo illustrazioni e disegni, non prove materiali.
Richard: Be', forse per vedere i fossili originali le toccherebbe andare al Museo di Nairobi, ma i calchi dei fossili, le copie esatte di quelli di Nairobi, li può vedere in qualunque grande museo si degni di visitare.
Wendy: Insomma, si può sapere perché è così aggressivo? Perché è così importante per lei che tutti credano a quello a cui crede lei?
Richard: Non sto parlando di credenze, ma di fatti. Le ho parlato di determinati fossili e ogni volta che le chiedo se è andata a vederli, lei elude la domanda e cambia discorso.
Wendy: Dovrebbero esserci valanghe di prove materiali, non solo qualcosa qui e là; invece no, di prove non ce ne sono.
Richard: Ho scelto i fossili di ominidi perché pensavo le sarebbero interessati soprattutto quelli, ma può trovare fossili analoghi in qualunque gruppo di vertebrati abbia voglia di nominare.
Wendy: Vorrei tornare su un punto: perché è così importante per lei che tutti credano all'evoluzione?
Richard: Non mi piace il termine «credenza». Preferisco chiedere alla gente di guardare le prove e infatti le sto chiedendo di guardarle... Vorrei che andasse in un museo, esaminasse i dati concreti e non credesse a chi le dice che non ci sono prove. Vada a vedere le prove.
Wendy [ride]: Sì, e le direi...
Richard: Non c'è niente da ridere. Insomma vada, vada davvero. Le ho parlato dei fossili di ominidi, ma può andare anche a esaminare le prove dell'evoluzione del cavallo, le prove dell'evoluzione dei primi mammiferi, le prove dell'evoluzione dei pesci, le prove della transizione da pesci ad anfibi e rettili terrestri. Troverà queste cose in qualsiasi museo serio. Apra gli occhi e guardi i fatti.
Wendy: Io invece le dico apra gli occhi e guardi quali comunità sono state edificate da coloro che credono in un Dio amorevole il quale ha creato ognuno di noi...
Può sembrare che in questa intervista io mi sia inutilmente incaponito a invitare la Wright ad andare in un museo e vedere le prove dell'evoluzione, ma l'ho fatto scientemente. Queste persone sono state addestrate a dire: «Non esistono fossili, mostratemi le prove, mostratemi anche un solo fossile», e lo dicono così spesso da finire per crederci. Così ho condotto un esperimento: ho menzionato a quella donna tre o quattro fossili e non le ho permesso di fare la furba ignorandoli. I risultati sono deprimenti, un classico esempio della tattica usata più di frequente dai negazionisti quando li si mette di fronte all'evidenza: ignorare le prove e ripetere il mantra: «Mostratemi i fossili. Dove sono i fossili? Non ci sono fossili. Vi chiedo solo di mostrarmi un fossile intermedio».
Altri si lasciano confondere dai nomi e dalla loro inevitabile tendenza a sottendere false divisioni che nella realtà non esistono. Tutti i fossili che potrebbero essere degli intermedi vengono sempre classificati come Homo o Australopithecus: nessuno è mai classificato come stadio intermedio; perciò non ci sono stadi intermedi. Ma, come ho spiegato, è una conseguenza delle convenzioni della nomenclatura zoologica, non un fatto del mondo reale. L'intermedio più perfetto che si possa immaginare continuerebbe a essere infilato a forza o nel genere Homo o nel genere Australopithecus. Anzi, con tutta probabilità sarebbe chiamato Homo da metà paleontologi e Australopithecus dall'altra metà. Purtroppo, invece di convenire che i fossili ambiguamente intermedi sono esattamente quelli che è previsto siano dalla teoria evoluzionistica, i paleontologi darebbero quasi sicuramente l'impressione contraria, scontrandosi con furia su questioni di terminologia.
E’ un po' come la distinzione tra adulto e minore prevista dalla legge. A fini giuridici, e anche per decidere se un giovane sia abbastanza vecchio da votare o arruolarsi nell'esercito, occorre operare una distinzione assoluta. In Gran Bretagna, l'età minima per votare fu portata nel 1969 da ventuno a diciotto anni (nel 1971 fu introdotta la stessa modifica negli Stati Uniti e nel 1975 in Italia). Adesso si sta parlando di portarla a sedici anni. Ma, qualunque sia l'età per votare stabilita dal codice civile, nessuno pensa seriamente che al rintoccare della mezzanotte del diciottesimo (o ventunesimo o sedicesimo) compleanno un ragazzo si trasformi in un'altra persona. Nessuno crede davvero che vi siano due tipi di persone, i bambini e gli adulti, senza «stadi intermedi». Ovviamente tutti capiamo che l'intero periodo della crescita è un solo, lungo esercizio di «intermediatezza». Si può forse dire che alcuni di noi non siano mai realmente cresciuti. Analogamente, l'evoluzione umana, dall'Australopithecus afarensis all'Homo sapiens, è consistita in una serie ininterrotta di genitori i quali mettevano al mondo figli che un tassonomista loro contemporaneo avrebbe sicuramente assegnato alla loro stessa specie. A posteriori e per motivi che non sono lontani da quelli giuridici, i moderni tassonornisti vogliono a tutti i costi applicare a ciascun fossile un'etichetta con su scritto qualcosa come Australopithecus o Homo. Le targhe dei musei non prevedono definizioni come «a metà strada tra Australopithecus africanus e Homo habilis». I negazionisti si appigliano a questa convenzione linguistica come se fosse la prova di una mancanza di intermedi nel mondo reale. Allora si potrebbe anche dire che non esiste l'adolescente perché ogni singola persona che guardiamo può essere o un adulto votante (sopra i diciott'anni) o un bambino non votante (sotto i diciott'anni). Sarebbe come dire che la necessità gìuridica di stabilire un'età minima per il voto politico dimostra che l'adolescenza non esiste.
Torniamo ai fossili. Se gli alfieri del creazionismo avessero ragione, Australopithecus sarebbe «solo una scimmia», sicché i suoi predecessori sarebbero irrilevanti per la ricerca degli «anelli mancanti». Nondimeno, converrà dare loro un'occhiata. Sono stati rinvenuti alcuni reperti piuttosto frammentari. Di Ardipithecus, vissuto 4‑5 milioni di anni fa, esistono soprattutto denti, però sono state trovate abbastanza ossa del cranio e dei piedi per ritenere che camminasse eretto, almeno secondo la maggior parte degli anatomisti che le hanno esaminate. Pressoché alle stesse conclusioni sono giunti lo scopritore di un fossile più antico, Orrorintugenensis (detto anche ((Uomo del Millennio»), e lo scopritore di Sahelanthropus tchadensis (detto anche «Toumai»,fig. 7.9).
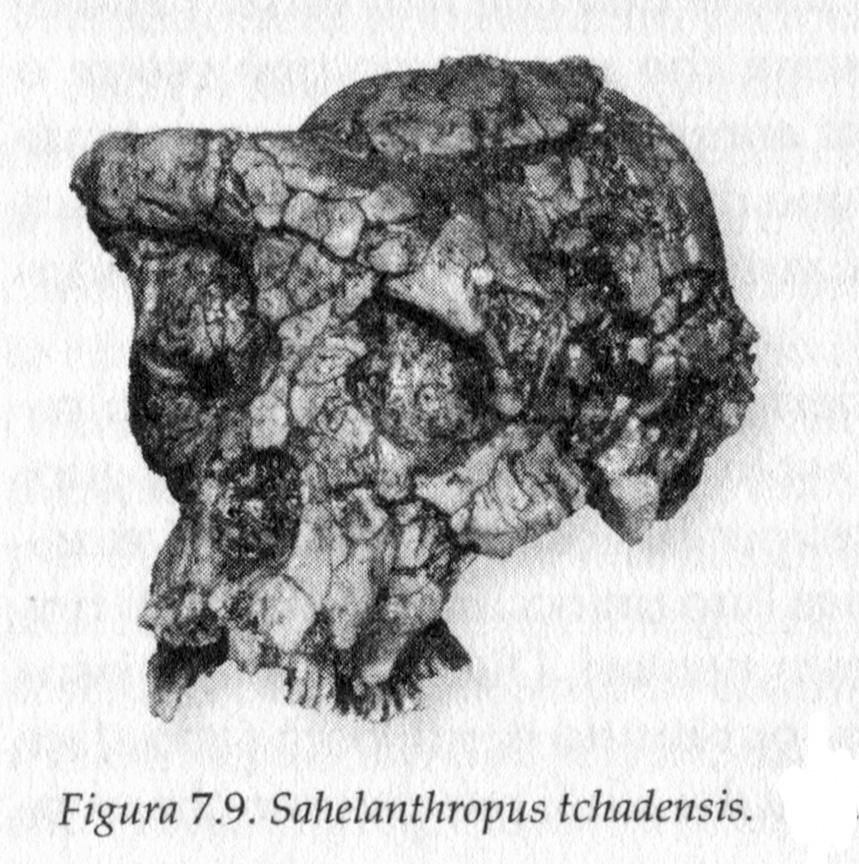
Sahelanthropus è notevole perché molto antico (sei milioni di anni, un'età vicina a quella dell'antenato che abbiamo in comune con gli scimpanzé) e perché è stato scoperto molto a ovest della Rift Valley (nel Ciad, dove «toumai» signifi «speranza di vita»). Altri paleoantropologi mettono in dubbio dichiarazioni degli scopritori, secondo i quali Orrorin e Sahelanthro pus sarebbero stati bipedi. E, come osserverebbe un cinico, tra coloro che mettono in dubbio quei fossili problematici vi sono gli scopritori degli altri...
Più di altre discipline scientifiche, la paleoantropologia è purtroppo funestata ‑ o è il caso di dire vivacizzata? ‑ dalle rivalità. Bisogna ammettere che la documentazione fossile capace di collegare la scimmia bipede Australopithecus con l'antenato (presumibilmente) quadrupede condiviso da uomo e scimpanzé è ancora scarsa. Non sappiamo come i nostri progenitori abbiano cominciato a camminare sulle zampe posteriori. Occorrono altri reperti. Ma rallegriamoci almeno della buona documentazione fossile che, diversamente da Darwin, abbiamo a nostra disposizione e che ci mostra la transizione evolutiva dall'Australopithecus, con il suo cervello da scimpanzé, all'Homo sapiens moderno, con il suo enorme cranio e il suo grande cervello.
In questo capitolo ho esibito immagini di crani, incoraggiandovi a confrontarli. Avrete forse notato, per esempio, il prognatismo o le arcate sopracciliari sporgenti di alcuni fossili. A volte la differenza è molto sottile, il che aiuta a capire quanto siano graduali le transizioni da un fossile a quello successivo
. Ora, però, vorrei introdurre una complicazione che si rivelerà a tutti gli effetti interessante. I cambiamenti che hanno luogo nell'arco della vita di un individuo a mano a mano che egli cresce, sono assai più cospicui dei cambiamenti che si rilevano confrontando adulti di generazioni successive.
Il cranio di uno scimpanzé prossimo alla nascita (fig. 7.10) è completamente diverso dal cranio di scimpanzé adulto (fig. 7.2, p. 161), e molto più simile a quello di un uomo (sia adulto sia bambino).
Le foto di uno scimpanzé cucciolo e di uno scimpanzé adulto (fig. 7.11) sono spesso usate per illustrare l'interessante idea che l'uomo mantenga caratteristiche infantili nell'età adulta (o l'idea non proprio identica che raggiunga la maturità sessuale avendo un corpo ancora infantile).
La foto del cucciolo mi pareva troppo bella per essere vera e l'ho mandata al mio collega Desmond Morris per sentire la sua opinione di esperto. Poteva essere un falso?, gli ho chiesto. Aveva mai visto un cucciolo di scimpanzé con un aspetto così umano? Morris non è convinto della schiena e delle spalle, ma è pienamente soddisfatto della testa. «E tipico degli scimpanzé avere una postura curva, mentre questo ha un collo umano meravigliosamente diritto, però, se prendi solo la testa, la foto è affidabile.» Sheila Lee, che ha diretto la ricerca iconografica per questo saggio, è risalita alla fonte originale della famosa foto, una spedizione in Congo organizzata dall'American Museum of Natural History negli anni 1909-1915. Gli animali erano morti quando furono fotografati e la Lee osserva che il fotografo, Herbert Lang, era anche tassidermista. Si sarebbe tentati di pensare che la postura stranamente umana del cucciolo di scimpanzé sia dovuta a una cattiva impagliatura; tuttavia, secondo il museo, Lang fotografava i suoi esemplari prima di imbalsamarli. Certo, si può correggere la postura di uno scimpanzé morto molto più di quanto non si possa fare con quella di uno scimpanzé vivo. La conclusione di Desmond Morris mi pare plausibile: la postura di tipo umano delle spalle del cucciolo può insospettire, ma la testa è degna di affidamento.
Se consideriamo autentica la testa, pur nutrendo qualche riserva sulle spalle, vediamo immediatamente come possa essere fuorviante un confronto tra crani fossili adulti. O, per dirla in maniera più costruttiva, la rilevante differenza tra testa adulta e testa infantile ci mostra con quanta facilità una caratteristica come il prognatismo possa imboccare la direzione giusta per diventare più ‑o meno ‑ umana. L'embriologia dello scimpanzé «sa» come produrre una testa di tipo umano, perché di fatto la produce per ogni scimpanzé nei suoi anni infantili. Sembra assai plausibile che Australopithecus, mentre evolveva attraverso vari stadi intermedi verso Homo sapiens e acquisiva un viso sempre meno prognato, abbia compiuto questa evoluzione tramite il metodo evidente di conservare caratteristiche infantili nell'età adulta (il processo chiamato neotenia e menzionato nel II capitolo). In ogni caso, molti cambiamenti evolutivi consistono in cambiamenti del ritmo al quale crescono certe parti del corpo rispetto ad altre: è la cosiddetta eterocronia o crescita eterocronica («a ritmi diversi»). Voglio dire, insomma, che il cambiamento evolutivo appare una bazzecola, una volta che si sia riconosciuto il fenomeno del cambiamento embriologico. Gli embrioni sono plasmati dalla crescita differenziale: le loro parti crescono a ritmi diversi. Nello scimpanzé cucciolo, le ossa delle mascelle e del muso si sviluppano in quelle dell'adulto a un ritmo relativamente rapido rispetto ad altre ossa del cranio. Riassumendo: tutti gli animali di tutte le specie cambiano, durante lo sviluppo embriologico, molto più di quanto non cambi la forma adulta tipica, generazione dopo generazione, nel corso delle ere geologiche.
Entrambi i capitoli sono un po’ farraginosi, ma sufficientemente espliciti per quanto riguarda l'intento di Dawkins di avallare il gradualismo e l’adattamentismo darwiniano in rapporto all’evoluzione delle forme viventi e dell’uomo.
I dati di cui oggi si dispone, però, sono poco o punto compatibili con tale intento.
Riguardo alla successione delle specie, nel secondo seminario su Darwin ho riassunto in questi termini ciò che oggi sostengono numerosi biologi evoluzionisti:
“La macroevoluzione sembra contrassegnata dal catastrofismo piuttosto che dal gradualismo.
Le prime tracce di vita (batteri fossili) risalgono a circa 3,4 miliardi di anni fa. Per almeno 2 miliardi di anni la vita è rappresentata solo da organismi unicellulari. Le prime testimonianze di animali pluricellulari risalgono alla fine del Precambiano. All’inizio del periodo Cambriano, in un intervallo di soli 10 milioni di anni, si realizza una vera e propria esplosione di diversificazione: compaiono tutti i principali gruppi di invertebrati con scheletro.
Circa 225 milioni di anni fa, nel periodo permiano, una metà delle famiglie di invertebrati marini di acque basse si estingue nell'arco di pochi milioni di anni. Nella scala dei grandi eventi della storia della vita questa "grande moria" è seconda solamente all'esplosione cambriana.
Un’altra grande estinzione avviene nel tardo Cretaceo e provoca la morte dei dinosauri, dei rettili volanti, dei rettili marini giganti, ecc., aprendo la via alla diversificazione dei mammiferi e alla loro radiazione planetaria.
In breve, il passaggio dal Precambriano al Paleozoico segna l'inizio dell'esplosione cambriana, il passaggio dal Paleozoico al Mesozoico l'estinzione permiana e quello fra il Mesozoico e il Cenozoico la seconda grande estinzione.
Questa ricostruzione, su cui concordano ormai gran parte dei paleontologi, è nettamente in contrasto con il gradualismo darwiniano.”
L'evoluzione delle forme viventi può essere rappresentata dalla figura seguente:
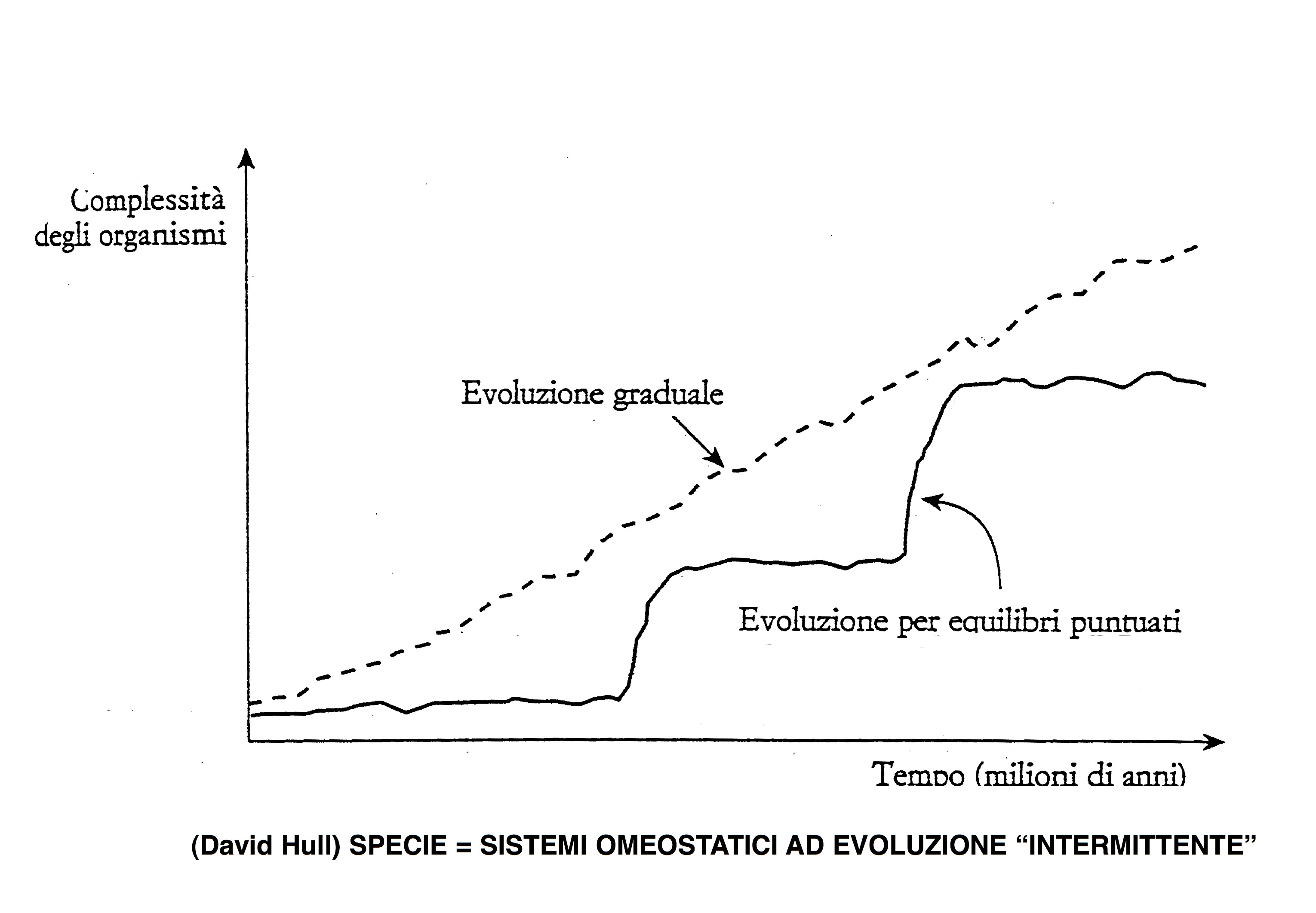
La curva tratteggiata corrisponde all'ipotesi gradualista di Darwin, che Dawkins ancora sostiene. La curva continua, però, sembra molto più fedele ai dati di cui oggi si dispone, che sono sinteticamente illustrati nella figura seguente:
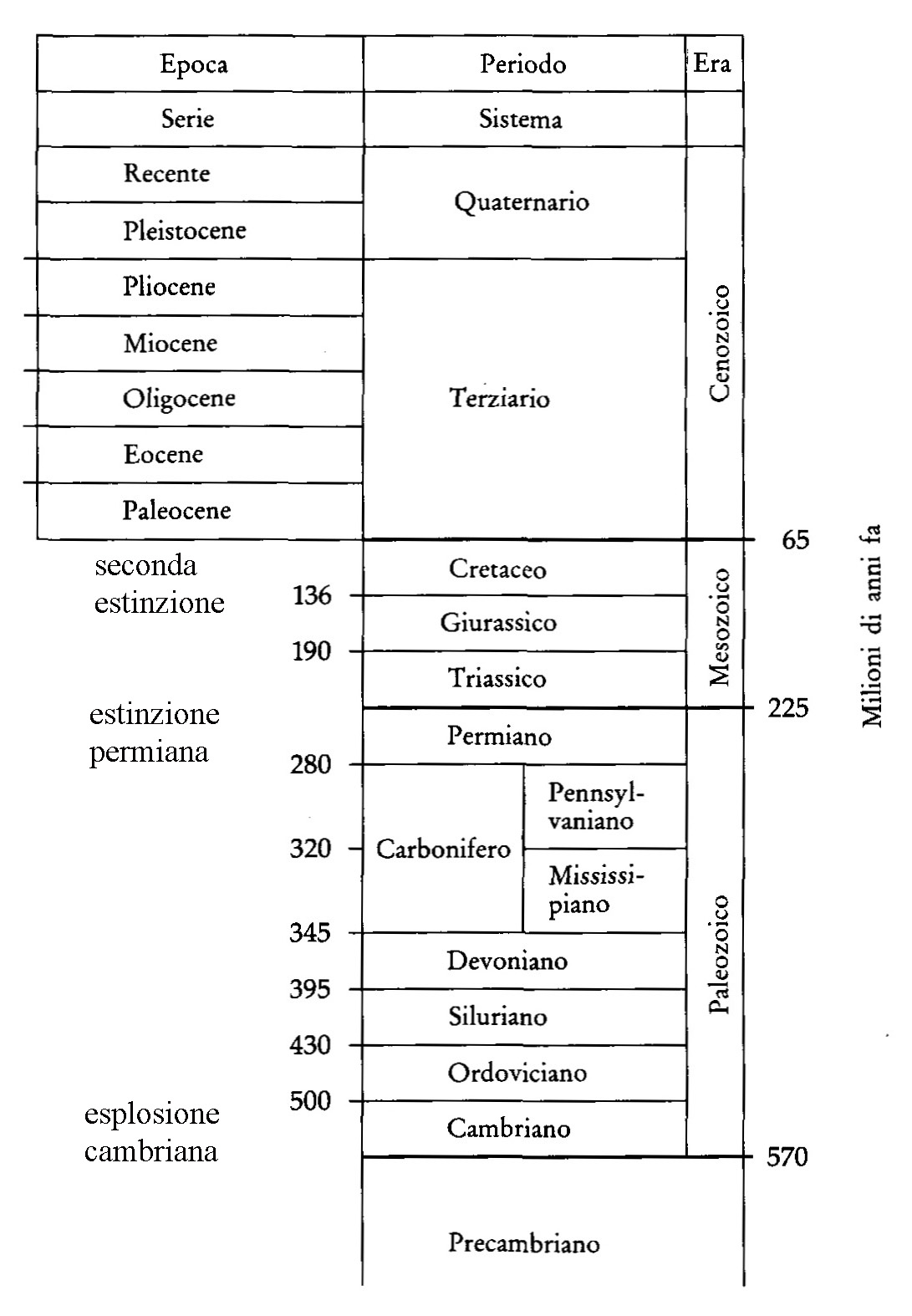
Ipotizzare come fa Dawkins che l’assenza di reperti fossili, che riguarda lunghissimi periodi geologici, possa essere attribuita o all’assenza di strutture scheletriche o a condizioni avverse alla fossilizzazione sembra veramente un arrampicarsi sugli specchi.
Per quanto riguarda l’evoluzione dell’uomo, la ricostruzione di Dawkins di un’evoluzione lineare, graduale e scalare, sembra ancora meno fondata. Nella quarta lettura darwiniana ho esposto nei seguenti termini il problema:
“I dati paleoantropologici (possono essere organizzati) in due modi diversi.
Il primo modo, adottato sin dagli anni 40 del secolo scorso dai neodarwinisti, consiste nell’ordinare i fossili in maniera tale che l’evoluzione degli ominidi risulti lineare, vale a dire caratterizzata da una crescita graduale del cervello e da un progressivo ingentilimento fisionomico. “Le forme pre-umane - scrive Pievani - si sostituiscono l'una all'altra gradualmente sotto la spinta della selezione naturale e di adattamenti progressivi: la forma che viene dopo, in virtù di un miglior adattamento, sostituisce la precedente”.
In questa ottica l’evoluzione degli ominidi è graduale e filetica, vale a dire dovuta soprattutto alle combinazioni e alle mutazioni genetiche che innescano lentissimi cambiamenti i quali infine esitano nella comparsa di una nuova specie. Essa, sorprendentemente, è anche lineare: si presume che le specie ominidi si succedano l’una l’altra (Homo abilis, Homo erectus, Home neandertalensis, Homo sapiens arcaico) con una progressione che culmina nella comparsa dell’Homo sapiens sapiens.
In tale progressione, ovviamente, per i neodarwinisti, non si dà alcun finalismo. Essa, però, suggestiona, implicando un tendere linearmente verso l’Uomo che non ha alcun riscontro nelle altre specie viventi, che sono ricche di varietà e di razze. “Mentre tutti i gruppi di mammiferi studiati dai paleontologi - scrive ancora Pievani - presentano un grafico evolutivo ad albero, con serie ordinate di specie che si diversificano progressivamente, l'evoluzione umana segue un andamento del tutto eterodosso, di tipo lineare e scalare.”
Da questo quadro affiora inesorabilmente la mitologia eroica dell’Uomo, che Pievani illustra nel modo seguente: “La sceneggiatura, accademica e popolare, riguardo alle origini dell'umanità divenne quella di una progressiva conquista della perfezione, con la frontiera del progresso umano che avanza verso l'intelligenza e la coscienza affrancandosi dalla condizione animale: una marcia a tappe serrate, dettata dall'espansione continua delle capacità cerebrali, letta sempre attraverso lo sguardo del vincitore, cioè della sola specie sopravvissuta al filtro della selezione naturale.”
Questa “mitologia” trova la massima espressione in Dobshansky, il quale nel 1942 afferma “il principio secondo cui non sarebbe stato mai possibile rinvenire due forme ominidi allo stesso livello temporale.”
Ad onore del vero, occorre aggiungere che i fondatori della Sintesi Moderna (come Theodosius Dobzhansky, George Gaylord Simpson ed Ernst Mayr) non erano ciechi di fronte al fatto che in natura esistono indizi di profonde discontinuità evolutive. Sembrava, però, loro difficile ammettere una discontinuità nell’evoluzione degli ominidi.
Il secondo modo di organizzare i dati paleantropologici comporta, invece, il superamento del gradualismo filetico in nome del fatto che i fossili paleantropologici sembrano alludere ad una storia del tutto diversa da quella “narrata” dai neodarwinisti. Nonché scalare e lineare, l’evoluzione degli ominidi appare come un cespuglio di specie che si diversificano coesistendo per lunghi periodi di tempo.
Rispetto allo schema lineare dei neodarwinisti che, come si è visto, comporta solo quattro specie che si succedono, il cespuglio comporta da 12 a 14 specie distinte divise in quattro generi: “i quattro gradini classici verso la perfezione di Homo sapiens - scrive Pievani - sono andati in frantumi e al loro posto troviamo una selva di ominidi ramificati e dispersi in un areale molto ampio.”
Anche l’Homo sapiens sapiens, comparso tra 150mila e 100mila anni fa è convissuto con due specie umane: l’Homo floresiensis e l’Home neandertalensis.
E’ evidente che questa ricostruzione cladica (a cespuglio) dell’ominazione fuoriesce dal paradigma del neodarwinismo. Essa porta ad ammettere non solo un’evoluzione non lineare e non graduale, ma qualche altro fattore oltre alla pressione genetica. Il nuovo paradigma, che corrobora l’evoluzionismo senza stravolgere le ipotesi di fondo di Darwin, identifica tale fattore nell’interazione tra geni e ambienti diversi. Valorizza, insomma, al massimo grado il ruolo della deriva genetica, che isolando geograficamente gruppi appartenenti alla stessa specie e impedendo loro di incrociarsi, amplifica la selezione naturale dei geni varianti.
La deriva genetica può promuovere salti evolutivi che non rientrano nel gradualismo darwiniano.”
I due modi di interpretare i dati paleoantrologici ssono rappresentati nelle figure seguenti:
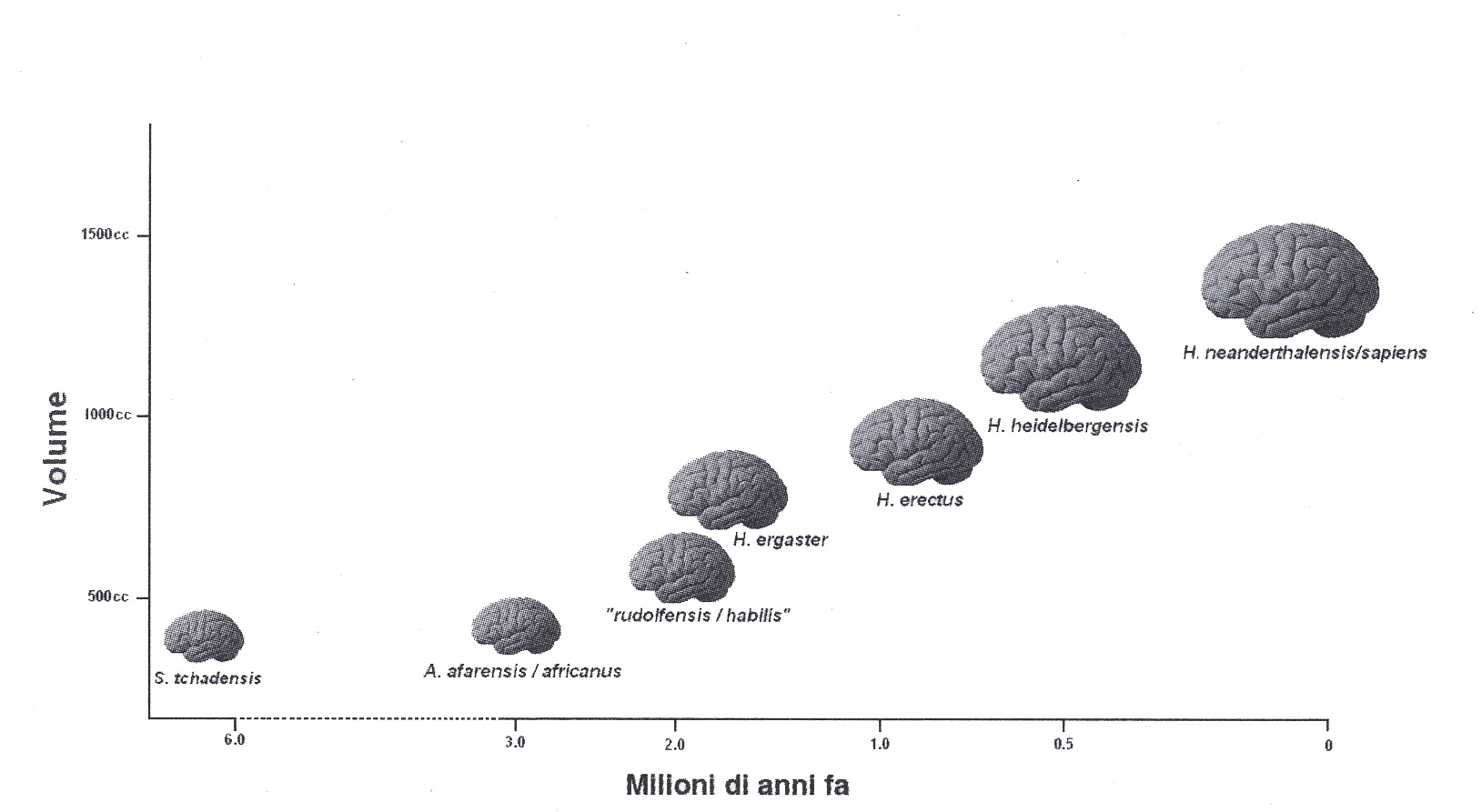
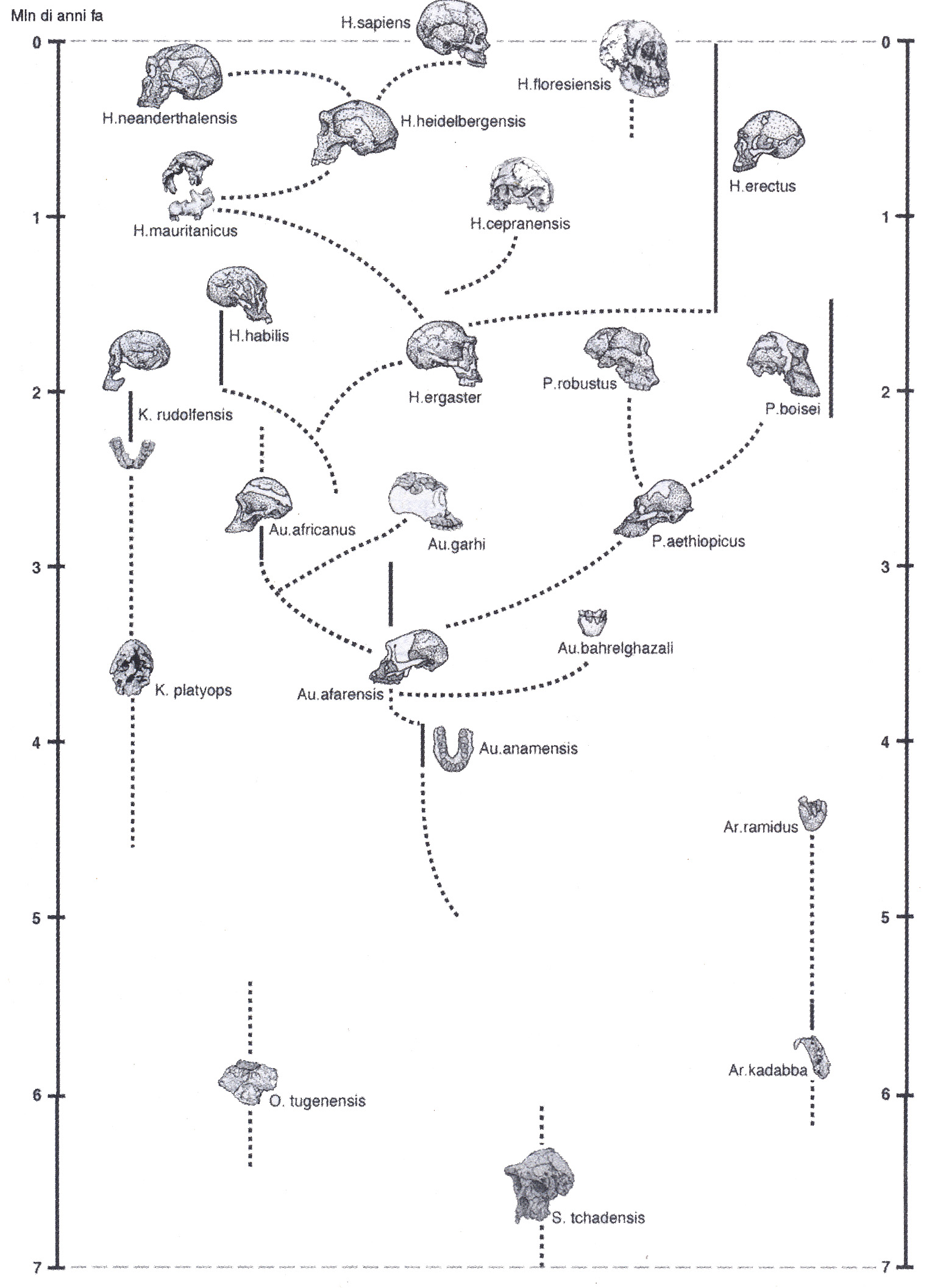
Ridurre i dati rappresentati nella seconda figura per ricavarne l'evoluzione lineare della prima, e confermare la linearità dell'ominazione, non sembra rispondere ad un criterio di onestà scientifica.
Cosa pensare del fatto che Dawkins trascura o manipola ad usum delphini dati che sono ormai noti a tutti i biologi evoluzionistici? L’unica ipotesi valida a riguardo è, forse, la motivazione che spinse Darwin ad accentuare un po’ troppo il ruolo della selezione naturale nella prima edizione de L’origine delle specie, vale a dire la preoccupazione di non fornire alcuno spiraglio al Creazionismo. Egli, però, nell’ultima edizione riconobbe che quella motivazione, per così dire, gli aveva preso un po’ la mano e fece ammenda.
In conseguenza del suo rigido e intollerante ateismo, che è rispettabile dato il contesto statunitense in cui opera, ma non dovrebbe interferire con l'alaborazione teorica, Dawkins non sembra minimamente intenzionato a seguire l’esempio del Maestro.
Non è una buona cosa, se si tiene conto che con il suo radicale neo-darwinismo, che è un po’ fuori tempo, egli non è riuscito a cooptare alcun Negazionista alla causa dell’evoluzionismo, mentre ha spinto Piattelli Palmarini e Fodor a scrivere un libro, esso stesso fuori tempo, vivacemente anti-neodarwinista, che, nonostante il dichiarato ateismo degli autori, è stato immediatamente “strumentalizzato” dai Negazionisti.
La scienza non deve avere alcun timore di esporre il fianco ad attacchi oscurantisti. Deve rimanere fedele a se stessa, vale a dire andare per la sua strada.